
You are using an out of date browser. It may not display this or other websites correctly.
You should upgrade or use an alternative browser.
You should upgrade or use an alternative browser.
CORONA Main Coronavirus thread
- Thread starter Heliobas Disciple
- Start date
-
- Tags
- coronavirus
Zoner
Veteran Member
Zoner
Veteran Member
Government & Media Pretending Massive Health Crisis Not Going On – Ed Dowd
By Greg Hunter On April 13, 2024 In Market Analysis 137 Comments
By Greg Hunter’s USAWatchdog.com (Saturday Night Post)
Former Wall Street money manager Ed Dowd is still a skillful number cruncher. His recently updated and wildly popular book “Cause Unknown: The Epidemic of Sudden Deaths in 2021, 2022 and 2023” has been correctly documenting the huge numbers of deaths and injuries caused by the CV19 bioweapon vax.
Many are waking up to this crime against humanity, but many remain in the dark because the government and Lying Legacy Media (LLM) continue to cover up the worst murder and disability fraud in world history.
Dowd says, “At this point, it’s overwhelming and has become almost comical. . . . This is asymmetric information. So, we have governments and media continuing to pretend a massive health crisis with chronic illness, deaths and disabilities is not going on.
The data would suggest otherwise. . . . The data we have made public is free, but some people want projections and decision-making ideas. These are things we might end up starting a business from. I would have never thought we could.
This is what asymmetric information does, and the government and the media are suppressing this information.”
A quick look at the overall casualties from the CV19 vax reveal an unparalleled medical disaster. Dowd explains, “I went before Senator Ron Johnson in February to talk about the ‘pandemic scorecard,’ which is abysmal.
Ever since the CV19 vaccine came on, we have had 1.1 million Americans die excessively, 4 million permanently disabled and another 28 million injured. It’s 33 million people who have been negatively affected now.
The question you have to ask is why are these institutions not screaming from the rooftops? I think the reason why is, it’s all because of the (deadly) vaccine. It’s all circular, and I think it’s a joke at this point.”
Is the worst over? The short answer from Dowd is No. Dowd contends, “Let’s just look at the disability data. We surged to a new high in June of 2023. We have not gone to a new high since. It kind of backfilled a little bit, but the last two months we have seen back-to-back increases. This is a called a plateauing effect. If it was all clear, I would like to see that number come down.
Unfortunately, it’s not. It can start to go back down, or it can have another consolidation and another spurt upward. The bad news is it is plateauing at a new high level. The good news is it has not gone up to a new level, but if it does, we have problems.”
One big problem Dowd has spotted is an explosion of cancers and, yes, you cannot get the truth about this either.
Dowd says, “The fact that people will not even say that cancers are on the rise is pretty comical to me. Doctors were reporting it anecdotally, and now we have the data to prove it. This is where we are.
In 2022, I said that ‘60,000 millennials died excessively between March of 2021 and February of 2022. That was a Vietnam War.’ That tweet went viral, and Reuters and AP fact checked me and said no, our experts say that’s not true. Now, even the establishment is saying there is excessive all-cause mortality. So, we are now in a stage where cancers are not rising. They are now denying that. The lies are just unreal.”
There is much more in the 36-minute interview.
Join Greg Hunter of USAWatchdog.com as he goes One-on-One with money manager and investment expert Ed Dowd, author of the recently updated book called “Cause Unknown: The Epidemic of Sudden Deaths in 2021, 2022 and 2023” for 4.13.24.
Government & Media Pretending Massive Health Crisis Not Going On – Ed Dowd | Greg Hunter’s USAWatchdog
Heliobas Disciple
TB Fanatic
(fair use applies)
Cambridge Study Finds That SARS-CoV-2 Omicron Infection Induces Immune Imprinting Even Prior To Vaccination
Nikhil Prasad Fact checked by:Thailand Medical News Team
Apr 15, 2024
The ongoing battle against the SARS-CoV-2 virus has been marked by a relentless struggle to keep pace with its evolution. Despite significant progress in vaccination efforts and global exposure leading to population-level immunity, new variants like Omicron continue to pose challenges to public health strategies. This COVID-19 News report delves into the intricate dynamics of immune responses, focusing on the phenomenon of immune imprinting and its implications, particularly in the context of the Omicron variant based on a study conducted by researchers from University of Cambridge-UK, IHVN-UK, Cambridge Addenbrookes hospital-UK and Cambridge University Hospitals NHS Foundation Trust-UK.
Understanding Immune Imprinting: Origins and Significance
Immune imprinting, also referred to as original antigenic sin (OAS), is a concept rooted in immunology that describes how the immune system's initial exposure to a pathogen influences subsequent responses to related strains. This phenomenon has far-reaching implications, as it can either enhance or hinder the body's ability to defend against evolving pathogens. Historical examples, such as the differential responses to the 2009 H1N1 influenza pandemic based on prior exposure, underscore the relevance of immune imprinting in infectious disease outcomes.
Examining Immune Responses to Omicron: Insights from a Nigerian Study
A comprehensive longitudinal study conducted in Nigeria provided a unique opportunity to dissect immune responses to SARS-CoV-2, particularly in the wake of the emergence of the Omicron variant. The study cohort, comprising individuals with diverse exposure histories, allowed for a nuanced exploration of immune imprinting effects and vaccine-induced immunity.
Findings from the Study: Imprinting Effects on Neutralization
Upon enrollment in January 2023, it was noted that 93% of participants had previous exposure to the ancestral Wu-1 virus, while 58% had been exposed to the Omicron virus, as indicated by the presence of anti-N IgG and anti-receptor binding domain (RBD) IgG antibodies specific to Wu-1 and Omicron. Interestingly, individuals without prior Omicron exposure exhibited significantly higher neutralization against Wu-1 compared to Omicron variants, aligning with expectations. However, participants who tested positive for anti-RBD Omicron IgG surprisingly showed a 2-fold decrease in serum neutralization titers against Omicron BA.1 compared to Wu-1. These findings strongly suggest the presence of imprinted immunity from ancestral pre-Omicron lineage viruses, with the older Wu-1 responses effectively dominating over more recent Omicron lineage infections, which may have been multiple in nature.
Moreover, among participants with previous Omicron exposure and evidence of imprinting, subsequent Omicron infections and Wu-1-based vaccination were linked to enhanced responses across variants, leading to the equalization of neutralization tite rs for both Wu-1 and Omicron variants. Despite this, the Omicron responses did not surpass ancestral responses, indicating the persistence of imprinting effects and only partial mitigation even with additional exposures and vaccinations.
While post-dose 1 vaccination elicited high-titer neutralization responses against ancestral and Omicron variants BA.1, BA.2, and BA.4 in nearly all participants, neutralization against the highly immune-evasive XBB recombinant variant remained notably lower. A second vaccine dose provided only a modest boost in neutralization against the XBB variant, highlighting the challenges posed by highly divergent variants in terms of immune response elicitation.
The study's findings revealed intriguing patterns in immune responses. Participants who had prior exposure to the ancestral Wu-1 virus demonstrated heightened neutralization against it compared to Omicron variants, highlighting the imprinting effects of previous infections. Even among individuals with recent Omicron exposure, neutralization titers against Omicron were notably lower than those against Wu-1, indicating the persistence of imprinting influences on immune responses.
Impact of Vaccination on Imprinting: Unraveling Vaccine-Induced Responses
The study also delved into the impact of vaccination on immune imprinting. While vaccines elicited robust immune responses, including effective neutralization against Omicron, there were nuances in the breadth and potency of neutralizing antibodies. Notably, subsequent vaccine doses provided limited additional boosting, suggesting that initial vaccination primed substantial immune memory but had limited capacity to overcome imprinting effects entirely.
Implications for Pandemic Response and Vaccination Strategies
The study's findings hold significant implications for pandemic response strategies and vaccination approaches. Understanding the nuances of immune imprinting is crucial for optimizing vaccine efficacy and tailoring vaccination strategies to address evolving viral variants. For unvaccinated or vulnerable populations, the use of Omicron-specific vaccines may offer a more comprehensive shield against the virus by bypassing imprinting effects from ancestral strains.
Discussion: Addressing Challenges and Future Considerations
The intricate interplay between prior exposure, vaccination, and immune responses to SARS-CoV-2 variants underscores the complexity of pandemic management. Adaptive vaccination strategies, ongoing surveillance, and continuous research are essential pillars in navigating the ever-changing landscape of viral evolution and immune dynamics. Furthermore, considerations for future zoonotic virus infections must take into account the potential impacts of immune imprinting on disease severity and outcomes.
Conclusion: Navigating Immune Imprinting in the Era of SARS-CoV-2 Variants
In conclusion, immune imprinting plays a pivotal role in shaping immune responses to SARS-CoV-2 variants, including the highly divergent Omicron strain. While vaccination remains a cornerstone of pandemic control efforts, strategies must evolve to address imprinting effects and ensure comprehensive population protection. Continued research, vigilance, and adaptability are paramount in confronting the challenges posed by emerging viral variants and safeguarding public health on a global scale.
The study findings were published on a preprint server and are currently being peer reviewed.
 www.researchsquare.com
www.researchsquare.com
Read Also:
 www.thailandmedical.news
www.thailandmedical.news
https://www.thailandmedical.news/ne...9-vaccine-causes-persistent-immune-imprinting
https://www.thailandmedical.news/ne...ance-of-sar-cov-2-caused-by-immune-imprinting
https://www.thailandmedical.news/ne...ologic-imprinting-from-previous-coronaviruses
https://www.thailandmedical.news/ne...al-strains-helps-via-negative-immune-imprinti
Cambridge Study Finds That SARS-CoV-2 Omicron Infection Induces Immune Imprinting Even Prior To Vaccination
Nikhil Prasad Fact checked by:Thailand Medical News Team
Apr 15, 2024
The ongoing battle against the SARS-CoV-2 virus has been marked by a relentless struggle to keep pace with its evolution. Despite significant progress in vaccination efforts and global exposure leading to population-level immunity, new variants like Omicron continue to pose challenges to public health strategies. This COVID-19 News report delves into the intricate dynamics of immune responses, focusing on the phenomenon of immune imprinting and its implications, particularly in the context of the Omicron variant based on a study conducted by researchers from University of Cambridge-UK, IHVN-UK, Cambridge Addenbrookes hospital-UK and Cambridge University Hospitals NHS Foundation Trust-UK.
Understanding Immune Imprinting: Origins and Significance
Immune imprinting, also referred to as original antigenic sin (OAS), is a concept rooted in immunology that describes how the immune system's initial exposure to a pathogen influences subsequent responses to related strains. This phenomenon has far-reaching implications, as it can either enhance or hinder the body's ability to defend against evolving pathogens. Historical examples, such as the differential responses to the 2009 H1N1 influenza pandemic based on prior exposure, underscore the relevance of immune imprinting in infectious disease outcomes.
Examining Immune Responses to Omicron: Insights from a Nigerian Study
A comprehensive longitudinal study conducted in Nigeria provided a unique opportunity to dissect immune responses to SARS-CoV-2, particularly in the wake of the emergence of the Omicron variant. The study cohort, comprising individuals with diverse exposure histories, allowed for a nuanced exploration of immune imprinting effects and vaccine-induced immunity.
Findings from the Study: Imprinting Effects on Neutralization
Upon enrollment in January 2023, it was noted that 93% of participants had previous exposure to the ancestral Wu-1 virus, while 58% had been exposed to the Omicron virus, as indicated by the presence of anti-N IgG and anti-receptor binding domain (RBD) IgG antibodies specific to Wu-1 and Omicron. Interestingly, individuals without prior Omicron exposure exhibited significantly higher neutralization against Wu-1 compared to Omicron variants, aligning with expectations. However, participants who tested positive for anti-RBD Omicron IgG surprisingly showed a 2-fold decrease in serum neutralization titers against Omicron BA.1 compared to Wu-1. These findings strongly suggest the presence of imprinted immunity from ancestral pre-Omicron lineage viruses, with the older Wu-1 responses effectively dominating over more recent Omicron lineage infections, which may have been multiple in nature.
Moreover, among participants with previous Omicron exposure and evidence of imprinting, subsequent Omicron infections and Wu-1-based vaccination were linked to enhanced responses across variants, leading to the equalization of neutralization tite rs for both Wu-1 and Omicron variants. Despite this, the Omicron responses did not surpass ancestral responses, indicating the persistence of imprinting effects and only partial mitigation even with additional exposures and vaccinations.
While post-dose 1 vaccination elicited high-titer neutralization responses against ancestral and Omicron variants BA.1, BA.2, and BA.4 in nearly all participants, neutralization against the highly immune-evasive XBB recombinant variant remained notably lower. A second vaccine dose provided only a modest boost in neutralization against the XBB variant, highlighting the challenges posed by highly divergent variants in terms of immune response elicitation.
The study's findings revealed intriguing patterns in immune responses. Participants who had prior exposure to the ancestral Wu-1 virus demonstrated heightened neutralization against it compared to Omicron variants, highlighting the imprinting effects of previous infections. Even among individuals with recent Omicron exposure, neutralization titers against Omicron were notably lower than those against Wu-1, indicating the persistence of imprinting influences on immune responses.
Impact of Vaccination on Imprinting: Unraveling Vaccine-Induced Responses
The study also delved into the impact of vaccination on immune imprinting. While vaccines elicited robust immune responses, including effective neutralization against Omicron, there were nuances in the breadth and potency of neutralizing antibodies. Notably, subsequent vaccine doses provided limited additional boosting, suggesting that initial vaccination primed substantial immune memory but had limited capacity to overcome imprinting effects entirely.
Implications for Pandemic Response and Vaccination Strategies
The study's findings hold significant implications for pandemic response strategies and vaccination approaches. Understanding the nuances of immune imprinting is crucial for optimizing vaccine efficacy and tailoring vaccination strategies to address evolving viral variants. For unvaccinated or vulnerable populations, the use of Omicron-specific vaccines may offer a more comprehensive shield against the virus by bypassing imprinting effects from ancestral strains.
Discussion: Addressing Challenges and Future Considerations
The intricate interplay between prior exposure, vaccination, and immune responses to SARS-CoV-2 variants underscores the complexity of pandemic management. Adaptive vaccination strategies, ongoing surveillance, and continuous research are essential pillars in navigating the ever-changing landscape of viral evolution and immune dynamics. Furthermore, considerations for future zoonotic virus infections must take into account the potential impacts of immune imprinting on disease severity and outcomes.
Conclusion: Navigating Immune Imprinting in the Era of SARS-CoV-2 Variants
In conclusion, immune imprinting plays a pivotal role in shaping immune responses to SARS-CoV-2 variants, including the highly divergent Omicron strain. While vaccination remains a cornerstone of pandemic control efforts, strategies must evolve to address imprinting effects and ensure comprehensive population protection. Continued research, vigilance, and adaptability are paramount in confronting the challenges posed by emerging viral variants and safeguarding public health on a global scale.
The study findings were published on a preprint server and are currently being peer reviewed.
Immune imprinting revealed by SARS-CoV-2 Omicron infection prior to vaccination
Immune imprinting or original antigenic sin (OAS) originally referred to a phenomenon of suboptimal immune response to a repeat exposure to a virus that was antigenically distinct from the original virus infection. OAS has been implicated in higher mortality in young people during the 2009-10 H1N...
www.researchsquare.com
Read Also:
COVID-19 News: Hybrid Immunity And Bivalent Boosters Have Little Effect On New Omicron Sub-Lineages BQ.1.1 And BA.2.75.2 Due To Immune Imprinting! - Thailand Medical News
COVID-19 News: Despite various governments, health authorities and mainstream media are trying to downplay the dire situation we are in with regards to the COVID-19 pandemic and are concealing COVID-19 statistics and also implications of long term catastrophic issues while making stupid claims...
www.thailandmedical.news
https://www.thailandmedical.news/ne...9-vaccine-causes-persistent-immune-imprinting
https://www.thailandmedical.news/ne...ance-of-sar-cov-2-caused-by-immune-imprinting
https://www.thailandmedical.news/ne...ologic-imprinting-from-previous-coronaviruses
https://www.thailandmedical.news/ne...al-strains-helps-via-negative-immune-imprinti
psychgirl
Has No Life - Lives on TB
…..I hope Geert comments on this finding.
Soon.
vector7
Dot Collector
This is absolutely INSANE.
"At least 15 federal agencies KNEW that gain of function research was being done in the WUHAN lab SINCE 2018"
"We know this because a whistleblower revealed it"
"Nobody in the administration told us anything"
"A brave marine a Lieutenant gave us that information"
"Or today we would still not know any of this"
So that means the government knew about COVID before 2021?
Never trust the deep state.
Because this was planned to steal an election.
RT 1min
View: https://twitter.com/imjdsharp/status/1779951184406593641
"At least 15 federal agencies KNEW that gain of function research was being done in the WUHAN lab SINCE 2018"
"We know this because a whistleblower revealed it"
"Nobody in the administration told us anything"
"A brave marine a Lieutenant gave us that information"
"Or today we would still not know any of this"
So that means the government knew about COVID before 2021?
Never trust the deep state.
Because this was planned to steal an election.
RT 1min
View: https://twitter.com/imjdsharp/status/1779951184406593641
psychgirl
Has No Life - Lives on TB
Annnd….nobody cares anymore except for us.This is absolutely INSANE.
"At least 15 federal agencies KNEW that gain of function research was being done in the WUHAN lab SINCE 2018"
"We know this because a whistleblower revealed it"
"Nobody in the administration told us anything"
"A brave marine a Lieutenant gave us that information"
"Or today we would still not know any of this"
So that means the government knew about COVID before 2021?
Never trust the deep state.
Because this was planned to steal an election.
RT 1min
View: https://twitter.com/imjdsharp/status/1779951184406593641
“Rug, meet sweeper” …

Tristan
Has No Life - Lives on TB
Annnd….nobody cares anymore except for us.
“Rug, meet sweeper” …
Most peoples' 'Horizon' is pretty short, both looking forward, and back.
Tristan
Has No Life - Lives on TB
This is absolutely INSANE.
"At least 15 federal agencies KNEW that gain of function research was being done in the WUHAN lab SINCE 2018"
"We know this because a whistleblower revealed it"
"Nobody in the administration told us anything"
"A brave marine a Lieutenant gave us that information"
"Or today we would still not know any of this"
So that means the government knew about COVID before 2021?
Never trust the deep state.
Because this was planned to steal an election.
RT 1min
View: https://twitter.com/imjdsharp/status/1779951184406593641
Not 'Insane'.
That's not the right word, at all...
psychgirl
Has No Life - Lives on TB
Evil work better?Not 'Insane'.
That's not the right word, at all...
I say both words
All I'll say is that it's been *extensively * studied in animals whose main/only purpose is reproduction. It's used routinely in racehorse stallions who get half a million dollars to breed ONE mare (and whose semen quality is routinely and frequently tested). It's used in dairy cattle, who, as a group, and probably the most closely monitored group of animals in the world, in terms of reproduction... herds get reports monthly on how many services per conception are required, and a change of one or two tenths is immediately followed up on. Ditto with the bulls who produce tens of thousands of units of semen yearly for artificial breeding.There are a group of blogger/substack writers who are very against ivermectin. I haven't posted about it because it seemed woo to me and possibly controlled opposition/disinformation. But it keeps coming up and there are some troubling real scientific studies on it done before covid ever came around. I am not trying to change anyone's mind, and because I know how controversial it is to even bring this up I'm just going to post links to these articles so you can decide for yourself and choose whether or not you want to read any further. The articles themselves are very long and image intensive and would take up 3 or 4 posts each to post so please do follow the links if you want to learn more. I want you to have all the information available so you can make an educated decision as you start or continue to take it. Sometimes it's a matter of balancing risk/benefit and most of us here are above child bearing age so this won't have an effect anyway, but good to know, especially if you are planning on giving it to your children. Who knows what Disease X is and when it's going to get here, ivermectin may be the best (only?) choice at that time. But make an educated decision, understand the risks; there are always risks with any drug you take (even aspirin, tylenol, etc). Why would ivermectin be any different?
"CLASTOGENIC" - 18 Studies Highlighting Ivermectin Induced DNA Breakage, Damage & Related Disorders
"CLASTOGENIC" - 18 Studies Highlighting Ivermectin Induced DNA Breakage and Damage
Definitions: A clastogen is a substance or agent that induces chromosomal breakage or structural changes in the DNA of cells. The term "clastogen" is derived from the Greek word "klastos," meaning "to break." Clastogens can cause damage to the genetic material, leading to the formation of...timtruth.substack.com
Cytotoxic, Mutagenic, Genotoxic & Clastogenic? Ivermectin Is A Menace
Ivermectin VS Sperm: 3 Disturbing Studies On Decreased Sperm Counts, Decreased Sperm Motility & Increased Abnormal Sperm Post-Ivermectin
Ivermectin VS Sperm: 3 Disturbing Studies On Decreased Sperm Counts, Decreased Sperm Motility & Increased % Abnormal Sperm Post-Ivermectin
Bad signs for Ivermectin's effects on male fertility!
Bad signs for Ivermectin's effects on male fertility
Ivermectin Is Cytotoxic & Genotoxic (Damaging To Cells And DNA) & Possibly Carcinogenic (Cancer Causing)
Ivermectin Is Cytotoxic & Genotoxic (Damaging To Cells And DNA) & Likely Carcinogenic (Cancer Causing)
Why were we told this was safe?
Why were we told this was safe?
Ivermectin Is So Toxic It Kills Most Mosquitoes That Feed On Its Users For 4 Days After?!
Ivermectin Is So Toxic It Kills Most Mosquitoes That Feed On Its Users For 4 Days After?!
Ivermectin is known to be an acutely toxic drug. According to one study, taken even at a standard dosage it kills most mosquitoes who feed on the Ivermectin user in the first 4 days. Much Higher Mortality And Fewer Children For Mosquitoes Feeding On Ivermectin Patients’ Blood
Human subjects became walking insecticide for days after standard Ivermectin dose
Ivermectin, WHO, UN, Merck, The World Bank & Kissinger's World Population Plan Of Action
Ivermectin, WHO, UN, Merck, The World Bank & Kissinger's World Population Plan Of Action
All signs point to genocide
All signs point to genocide
The Branding and the Re-Branding of the Wonderful Mectizan (Ivermectin)
The Branding and the Re-Branding of the Wonderful Mectizan (Ivermectin)
"semi-synthetic anti parasitic", anti viral, anti-helmintic, anti cancer, washes dishes, mows lawns, and if you stack it with Pepcid AC, plays Beethovensagehana.substack.com
"semi-synthetic anti parasitic", anti viral, anti-helmintic, anti cancer, washes dishes, mows lawns, and if you stack it with Pepcid AC, plays Beethoven
If ANY of the above scary baloney was true.. We'd know it!
However, it's been pretty obvious that the Covid vax (and probably the virus, as well, to a lesser extent) has ALL the above problems.
Also... "humans become a walking insecticide for days" after a dose of ivermectin...duh! That's how it is designed to work! It is routinely used to prevent louse infection in various livestock for up to 3 weeks.
Summerthyme
Last edited:
John Deere Girl
Veteran Member
Also, I'd think a lot of farmers would be showing side effects from wearing it when applying pour on for cattle and pigs when the livestock weren't exactly cooperative. I know I've worn my fair share.All I'll say is that it's been *extensively * studied in animals whose main/only purpose is reproduction. It's used routinely in racehorse stallions who get half a million dollars to breed ONE mare (and whose seven quality is routinely and frequently tested). It's used in dairy cattle, who, as a group, and probably the most closely monitored group of animals in the world, in terms of reproduction... herds get reports monthly on how many services per conception are required, and a change of one or two tenths is immediately followed up on. Ditto with the bulls who produce tens of thousands of units of seven yearly for artificial breeding.
If ANY of the above scare baloney was true.. We'd know it!
However, it's been pretty obvious that the Covid vax (and probably the virus, as well, to a lesser extent) has ALL the above problems.
Also... "humans become a walking insecticide for days" after a dose of ivermectin...duh! That's how it 8s designed to work! It is routinely used to prevent louse infection in various livestock for up to 3 weeks.
Summerthyme
Could you provide a link, please?Amazon sells the portable oxygen machines and the big ones for home use. The one we bought for my DH is exactly like the one my FIL has. No prescription required. We've had it for three years with no issues.
Summerthyme
Profit of Doom
Dismember
Consider how long the small tank would last. My wife was generally at 5 litres per minute.I refer you to research the web for purity..I did they are not identical.
plus if are truly that sick would you really want to truck to yourself garage to breath oxygen and change fittings or try to get that heavy tank in your house..if you are needing oxygen you will very very sick..
I figure having the cans of oxygen will be enough to get you to your doctor's office without dragging your welding tank..our tank is huge
In addition not everyone has welding tanks just sitting around..
Profit of Doom
Dismember
We never got billed for oxygen, please consider that cost vs. the investment, and logistics for availability and delivery.I went to Amazon and researched them..
It looks like the least expensive one with shipping and travel bag and required add-ons will cost north of $800..
That could be too expensive for most people..but it is less expensive than many of the name brand ones..
I was going to post a thread, and probably will..but..
I'm going to have to increase my prices on most products.... the honey for elderberry syrup has doubled in price! But I have a couple fresh batches left for $12 a bottle... it will be going up to $15 after these are gone.
Elderberry works (in many studies) at least as well as Tamiflu, and AFAIK, no one has ever committed suicide after taking elderberry! PM me if you are interested.
Summerthyme
I'm going to have to increase my prices on most products.... the honey for elderberry syrup has doubled in price! But I have a couple fresh batches left for $12 a bottle... it will be going up to $15 after these are gone.
Elderberry works (in many studies) at least as well as Tamiflu, and AFAIK, no one has ever committed suicide after taking elderberry! PM me if you are interested.
Summerthyme
John Deere Girl
Veteran Member
I will but it will be a bit later. Remind me if I forget please!Could you provide a link, please?
Summerthyme
Thank you!
Summerthyme
Summerthyme
Heliobas Disciple
TB Fanatic
Modified RNA in COVID Vaccines May Contribute to Cancer Development: Review
In a review published on April 5, researchers argue that modification to mRNA causes immune suppression that may contribute to cancer development.
 www.theepochtimes.com
www.theepochtimes.com
Modified RNA in COVID Vaccines May Contribute to Cancer Development: Review
In a review published on April 5, researchers argue that modification to mRNA causes immune suppression that may contribute to cancer development.
By Marina Zhang,
4/15/2024
The mRNA used in the COVID-19 vaccines differs from naturally occurring mRNA, which has been modified to prevent immune degradation when injected. In a review published on April 5, researchers argue that modification—specifically, N1-methyl-pseudouridine modification—to mRNA causes immune suppression that may contribute to cancer development.
Uridine is a key component of mRNA. However, when uridine is injected into the body, the immune system rapidly degrades it.
Therefore, scientists at Pfizer and Moderna modified the uridine to N1-methyl-pseudouridine (m1-psi) so it could last longer.
However, research suggests this modification may reduce immune responses. “Within the framework of COVID-19 vaccination, this inhibition ensures an appropriate spike protein synthesis and a reduced immune activation,” the authors wrote in the study’s abstract.
The authors are concerned the modification may promote cancer in susceptible individuals.
“We suggest that future clinical trials for cancers or infectious diseases should not use mRNA vaccines with a 100% m1? (m1-psi) modification, but rather ones with the lower percentage of m1? modification to avoid immune suppression,” they wrote.
Modified vs. Natural RNA
Modified and natural RNA stimulate different responses in the body. Modified RNA tends to produce more aberrant proteins, potentially contributing to cell genome instability.
Most importantly, modified RNA induces a more muted response in the body than natural RNA, which may have broad implications for the body’s ability to fight other infections and cancers.
The authors cited studies finding that natural RNA tended to stimulate the activity of type-1 interferon—a key anti-tumor substance—and other immune chemicals. Contrastingly, modified RNA stimulates a milder response and is associated with immune chemicals that promote tolerance of foreign RNA injections.
A critical piece of evidence the review authors cited comes from a mice study in Thailand.
In the study, researchers injected natural mRNA and 100 percent modified RNA into two different groups of mice with melanoma.
The Thai researchers found that when injected with natural RNA, the body had a more robust immune response than when injected with modified RNA.
Furthermore, the prognosis of the melanoma mice also differed.
The survival rate in the mice group without modified RNA was 100 percent. Conversely, in the group with the modified uridine, only half of the mice survived.
In their abstract, the Thai researchers wrote that an mRNA vaccine induces type-1 interferon production and downstream signals crucial for controlling tumor growth and metastasis.
The review authors interpreted the findings to mean that adding 100 percent of “[m1-psi] to the mRNA vaccine in a melanoma model stimulated cancer growth and metastasis, while non-modified mRNA vaccines induced opposite results.”
They added that the study potentially shows that complete mRNA modification reduced survival in recipients.
The Cancer Debate
However, Tanapat Palaga, who has a doctorate in microbiology and immunology, is a professor of microbiology at Chulalongkorn University in Bangkok, Thailand, and is the senior author of the Thai study, told The Epoch Times in an email that the review took his team’s “results out of the context.”
While Mr. Palaga agreed that the unmodified RNA is associated with “robust anti-tumor immunity,” he added that their study “did not indicate, conclude or suggest that modified mRNA promotes tumor formation.
“Modified RNA ... just simply did not induce IFN type I production,” he wrote.
The review authors were careful to highlight that they are not suggesting that modified RNA causes cancer but that its effects may lead to an environment that aids in the development of cancers.
“Those who do not read in depth will be quick to say that we are AFFIRMING that mRNA vaccines cause cancer,” the review authors wrote to The Epoch Times in an email, pointing to a passage in their paper stating, “It is important to clarify here that mRNA vaccines do not cause cancer; but they could stimulate its development ... We are more concerned with experimental and clinical data with regard to the latter.”
Dr. Tian Xia, a professor in the Division of NanoMedicine at the University of California–Los Angeles, told The Epoch Times that the basis of m1-psi is to reduce immunotoxicity “not to suppress the innate and adaptive immunity,” and the cancer-causing conclusion doesn’t have “strong scientific support.”
Reduce Modified RNA Use
The review authors suggested that mRNA therapeutics should include a “lower percentage” of modified RNA in the future.
They also wrote that they do not discourage the use of mRNA injections in cancer treatment, given that natural or unmodified RNA reduces tumor growth, improves the efficiency of immune responses, and may potentially increase survival.
The authors told The Epoch Times that with COVID-19 mRNA vaccines, the scientists “only focused on maximizing the production of the spike protein” without considering other downstream effects.
“We have to do a deep reflection here: if you lower the [percentage] of modification, you have a less effective vaccine against SARS-CoV-2,” the authors wrote, but at the same time, there may be fewer unintended adverse effects.
Raquel Valdes Angues, a senior research associate at Oregon Health & Science University, told The Epoch Times that she and her colleagues welcome the review “addressing the potential implications of using COVID-19 [m1-psi]-modified mRNA vaccines on cancer progression and metastasis.”
She highlighted that modified RNA has been shown to impede interferon signaling, and given its complex role in tumor biology, “it becomes imperative to exercise caution when integrating modified [m1-psi] mRNAs for therapeutic use” in live animals and humans.
“These considerations warrant thorough investigation and thoughtful deliberation in the pursuit of mRNA-based therapeutics,” she said.
Heliobas Disciple
TB Fanatic
New vaccine strategy may mean the end of the line for endless boosters
Scientists at UC Riverside have demonstrated a new, RNA-based vaccine strategy that is effective against any strain of a virus and can be used safely even by babies or the immunocompromised.
New vaccine strategy may mean the end of the line for endless boosters
by University of California - Riverside
April 15, 2024
Scientists at UC Riverside have demonstrated a new, RNA-based vaccine strategy that is effective against any strain of a virus and can be used safely even by babies or the immunocompromised.
Every year, researchers try to predict the four influenza strains that are most likely to be prevalent during the upcoming flu season. And every year, people line up to get their updated vaccine, hoping the researchers formulated the shot correctly.
The same is true of COVID vaccines, which have been reformulated to target sub-variants of the most prevalent strains circulating in the U.S.
This new strategy would eliminate the need to create all these different shots because it targets a part of the viral genome that is common to all strains of a virus. The vaccine, how it works, and a demonstration of its efficacy in mice is described in a paper published in the Proceedings of the National Academy of Sciences.
"What I want to emphasize about this vaccine strategy is that it is broad," said UCR virologist and paper author Rong Hai. "It is broadly applicable to any number of viruses, broadly effective against any variant of a virus, and safe for a broad spectrum of people. This could be the universal vaccine that we have been looking for."
Traditionally, vaccines contain either a dead or modified, live version of a virus. The body's immune system recognizes a protein in the virus and mounts an immune response. This response produces T-cells that attack the virus and stop it from spreading. It also produces "memory" B-cells that train your immune system to protect you from future attacks.
The new vaccine also uses a live, modified version of a virus. However, it does not rely on the vaccinated body having this traditional immune response or immune active proteins—which is the reason it can be used by babies whose immune systems are underdeveloped, or people suffering from a disease that overtaxes their immune system. Instead, this relies on small, silencing RNA molecules.
"A host—a person, a mouse, anyone infected—will produce small interfering RNAs as an immune response to viral infection. These RNAi then knock down the virus," said Shouwei Ding, distinguished professor of microbiology at UCR, and lead paper author.
The reason viruses successfully cause disease is because they produce proteins that block a host's RNAi response. "If we make a mutant virus that cannot produce the protein to suppress our RNAi, we can weaken the virus. It can replicate to some level, but then loses the battle to the host RNAi response," Ding said. "A virus weakened in this way can be used as a vaccine for boosting our RNAi immune system."
When the researchers tested this strategy with a mouse virus called Nodamura, they did it with mutant mice lacking T and B cells. With one vaccine injection, they found the mice were protected from a lethal dose of the unmodified virus for at least 90 days. Note that some studies show nine mouse days are roughly equivalent to one human year.
There are few vaccines suitable for use in babies younger than six months old. However, even newborn mice produce small RNAi molecules, which is why the vaccine protected them as well. UC Riverside has now been issued a US patent on this RNAi vaccine technology.
In 2013, the same research team published a paper showing that flu infections also induce us to produce RNAi molecules. "That's why our next step is to use this same concept to generate a flu vaccine, so infants can be protected. If we are successful, they'll no longer have to depend on their mothers' antibodies," Ding said.
Their flu vaccine will also likely be delivered in the form of a spray, as many people have an aversion to needles. "Respiratory infections move through the nose, so a spray might be an easier delivery system," Hai said.
Additionally, the researchers say there is little chance of a virus mutating to avoid this vaccination strategy. "Viruses may mutate in regions not targeted by traditional vaccines. However, we are targeting their whole genome with thousands of small RNAs. They cannot escape this," Hai said.
Ultimately, the researchers believe they can 'cut and paste' this strategy to make a one-and-done vaccine for any number of viruses.
"There are several well-known human pathogens; dengue, SARS, COVID. They all have similar viral functions," Ding said. "This should be applicable to these viruses in an easy transfer of knowledge."
More information: Hai, Rong et al, Live-attenuated virus vaccine defective in RNAi suppression induces rapid protection in neonatal and adult mice lacking mature B and T cells, Proceedings of the National Academy of Sciences (2024). DOI: 10.1073/pnas.2321170121. doi.org/10.1073/pnas.2321170121
Journal information: Proceedings of the National Academy of Sciences
Provided by University of California - Riverside
Heliobas Disciple
TB Fanatic
(fair use applies)
Norwegian Study Shows That Cardiac Troponin T Can Be Used As A Biomaker In Moderate-To-Severe COVID-19 To Determine Potential Heart Issues
Nikhil Prasad Fact checked by:Thailand Medical News Team
Apr 16, 2024
The outbreak of COVID-19 has sparked numerous investigations into its multifaceted impacts on human health. Among the myriad of effects associated with this viral infection, cardiovascular implications have garnered significant attention. One area of interest is the behavior of cardiac troponin T (cTnT), a biomarker indicative of myocardial injury. Elevated cTnT levels have been observed in patients with COVID-19, raising questions about potential cardiac involvement and long-term consequences.
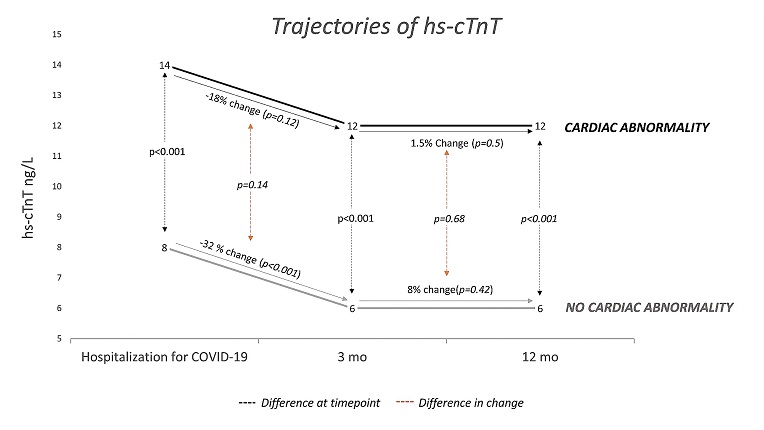
Trajectory of cardiac troponin T following moderate-to-severe COVID-19 and the association with cardiac abnormalities
Trajectory of high sensitivity cardiac troponin T (hs-cTnT) from peak concentration during hospitalization for COVID-19 to 3 and 12 months after discharge. Stratified by the presence of cardiac abnormality after 3 months and presented as geometric mean. Patients with hs-cTnT available at all time points (n?=?125).
Background and Objectives of the Study
This COVID-19 News report covers a recent study by researchers from multiple institutions in Norway, including Akershus University Hospital, the University of Oslo, and others, who collaborated to delve into this complex interplay between COVID-19 and cardiac troponin T. Their primary objective was to understand how high-sensitive cardiac troponin T (hs-cTnT) concentrations correlate with cardiac dysfunction and structural abnormalities post-hospitalization for COVID-19. Additionally, they aimed to evaluate hs-cTnT's efficacy in ruling out cardiac pathology.
Methodology: Examining Patients and Assessing Cardiac Health
The study involved a cohort of 189 patients hospitalized with COVID-19. These individuals underwent hs-cTnT measurements during their hospital stay and at 3- and 12-month follow-up visits. Additionally, they underwent echocardiographic studies, and a subset received cardiovascular magnetic resonance imaging (CMR) after six months. Cardiac abnormalities were defined based on various parameters, including left ventricular hypertrophy or dysfunction, right ventricular dysfunction, and CMR late gadolinium findings.
Findings and Interpretations: Unraveling the Insights
The study's findings provided several key insights into the relationship between hs-cTnT and cardiac health post-COVID-19:
- Association with Cardiac Abnormalities: Higher hs-cTnT concentrations during convalescence were associated with cardiac pathology, indicating a potential link between COVID-19 severity and lasting cardiac effects.
- Decrease in hs-cTnT Levels: Interestingly, hs-cTnT levels decreased significantly from hospitalization to three months post-discharge, especially in patients treated in intensive care units (ICUs). This suggests a transient nature of hs-cTnT elevation related to acute illness.
- Role in Ruling Out Cardiac Pathology: Hs-cTnT levels below a certain threshold (<5 ng/L) showed a high negative predictive value for ruling out cardiac abnormalities after COVID-19, providing a potentially useful tool in clinical assessments.
- Persistent Dyspnea and Cardiac Abnormalities: The study also explored the relationship between persistent dyspnea (a symptom in Post-Acute COVID Syndrome) and cardiac abnormalities, finding a significant association between hs-cTnT levels and the presence of cardiac pathology in dyspneic patients.
Contextualizing the Findings and Implications
These findings contribute significantly to our understanding of post-COVID-19 cardiac health. They suggest that while elevated hs-cTnT during acute illness may reflect severity and acute myocardial injury, hs-cTnT levels during convalescence could serve as markers for lasting cardiac effects. Moreover, the study underscores the importance of hs-cTnT in risk stratification and clinical decision-making regarding post-COVID-19 cardiac assessments.
Strengths, Limitations, and Future Directions
The study's strengths lie in its prospective design, inclusion of unselected patients, and comprehensive cardiac evaluations. However, limitations such as survival bias and lack of pre-COVID cardiac health data highlight areas for further research. Future studies could explore longitudinal hs-cTnT trends beyond 12 months and investigate potential interventions based on hs-cTnT levels in post-COVID-19 cardiac management.
Conclusion: Insights and Clinical Implications
In conclusion, the Norwegian study sheds light on the nuanced relationship between hs-cTnT and cardiac health post-COVID-19. It emphasizes the dynamic nature of hs-cTnT levels, their role in predicting cardiac pathology, and their potential utility in clinical decision-making for post-COVID-19 patients. These findings pave the way for personalized approaches to cardiac care in the aftermath of COVID-19.
The study findings were published in the peer reviewed journal: BMC Cardiovascular Disorders (Springer).

 bmccardiovascdisord.biomedcentral.com
bmccardiovascdisord.biomedcentral.com
Norwegian Study Shows That Cardiac Troponin T Can Be Used As A Biomaker In Moderate-To-Severe COVID-19 To Determine Potential Heart Issues
Nikhil Prasad Fact checked by:Thailand Medical News Team
Apr 16, 2024
The outbreak of COVID-19 has sparked numerous investigations into its multifaceted impacts on human health. Among the myriad of effects associated with this viral infection, cardiovascular implications have garnered significant attention. One area of interest is the behavior of cardiac troponin T (cTnT), a biomarker indicative of myocardial injury. Elevated cTnT levels have been observed in patients with COVID-19, raising questions about potential cardiac involvement and long-term consequences.
Trajectory of cardiac troponin T following moderate-to-severe COVID-19 and the association with cardiac abnormalities
Trajectory of high sensitivity cardiac troponin T (hs-cTnT) from peak concentration during hospitalization for COVID-19 to 3 and 12 months after discharge. Stratified by the presence of cardiac abnormality after 3 months and presented as geometric mean. Patients with hs-cTnT available at all time points (n?=?125).
Background and Objectives of the Study
This COVID-19 News report covers a recent study by researchers from multiple institutions in Norway, including Akershus University Hospital, the University of Oslo, and others, who collaborated to delve into this complex interplay between COVID-19 and cardiac troponin T. Their primary objective was to understand how high-sensitive cardiac troponin T (hs-cTnT) concentrations correlate with cardiac dysfunction and structural abnormalities post-hospitalization for COVID-19. Additionally, they aimed to evaluate hs-cTnT's efficacy in ruling out cardiac pathology.
Methodology: Examining Patients and Assessing Cardiac Health
The study involved a cohort of 189 patients hospitalized with COVID-19. These individuals underwent hs-cTnT measurements during their hospital stay and at 3- and 12-month follow-up visits. Additionally, they underwent echocardiographic studies, and a subset received cardiovascular magnetic resonance imaging (CMR) after six months. Cardiac abnormalities were defined based on various parameters, including left ventricular hypertrophy or dysfunction, right ventricular dysfunction, and CMR late gadolinium findings.
Findings and Interpretations: Unraveling the Insights
The study's findings provided several key insights into the relationship between hs-cTnT and cardiac health post-COVID-19:
- Association with Cardiac Abnormalities: Higher hs-cTnT concentrations during convalescence were associated with cardiac pathology, indicating a potential link between COVID-19 severity and lasting cardiac effects.
- Decrease in hs-cTnT Levels: Interestingly, hs-cTnT levels decreased significantly from hospitalization to three months post-discharge, especially in patients treated in intensive care units (ICUs). This suggests a transient nature of hs-cTnT elevation related to acute illness.
- Role in Ruling Out Cardiac Pathology: Hs-cTnT levels below a certain threshold (<5 ng/L) showed a high negative predictive value for ruling out cardiac abnormalities after COVID-19, providing a potentially useful tool in clinical assessments.
- Persistent Dyspnea and Cardiac Abnormalities: The study also explored the relationship between persistent dyspnea (a symptom in Post-Acute COVID Syndrome) and cardiac abnormalities, finding a significant association between hs-cTnT levels and the presence of cardiac pathology in dyspneic patients.
Contextualizing the Findings and Implications
These findings contribute significantly to our understanding of post-COVID-19 cardiac health. They suggest that while elevated hs-cTnT during acute illness may reflect severity and acute myocardial injury, hs-cTnT levels during convalescence could serve as markers for lasting cardiac effects. Moreover, the study underscores the importance of hs-cTnT in risk stratification and clinical decision-making regarding post-COVID-19 cardiac assessments.
Strengths, Limitations, and Future Directions
The study's strengths lie in its prospective design, inclusion of unselected patients, and comprehensive cardiac evaluations. However, limitations such as survival bias and lack of pre-COVID cardiac health data highlight areas for further research. Future studies could explore longitudinal hs-cTnT trends beyond 12 months and investigate potential interventions based on hs-cTnT levels in post-COVID-19 cardiac management.
Conclusion: Insights and Clinical Implications
In conclusion, the Norwegian study sheds light on the nuanced relationship between hs-cTnT and cardiac health post-COVID-19. It emphasizes the dynamic nature of hs-cTnT levels, their role in predicting cardiac pathology, and their potential utility in clinical decision-making for post-COVID-19 patients. These findings pave the way for personalized approaches to cardiac care in the aftermath of COVID-19.
The study findings were published in the peer reviewed journal: BMC Cardiovascular Disorders (Springer).
Trajectory of cardiac troponin T following moderate-to-severe COVID-19 and the association with cardiac abnormalities - BMC Cardiovascular Disorders
Background COVID-19 has been associated with cardiac troponin T (cTnT) elevations and changes in cardiac structure and function, but the link between cardiac dysfunction and high-sensitive cardiac troponin T (hs-cTnT) in the acute and convalescent phase is unclear. Objective To assess whether...
bmccardiovascdisord.biomedcentral.com
Heliobas Disciple
TB Fanatic
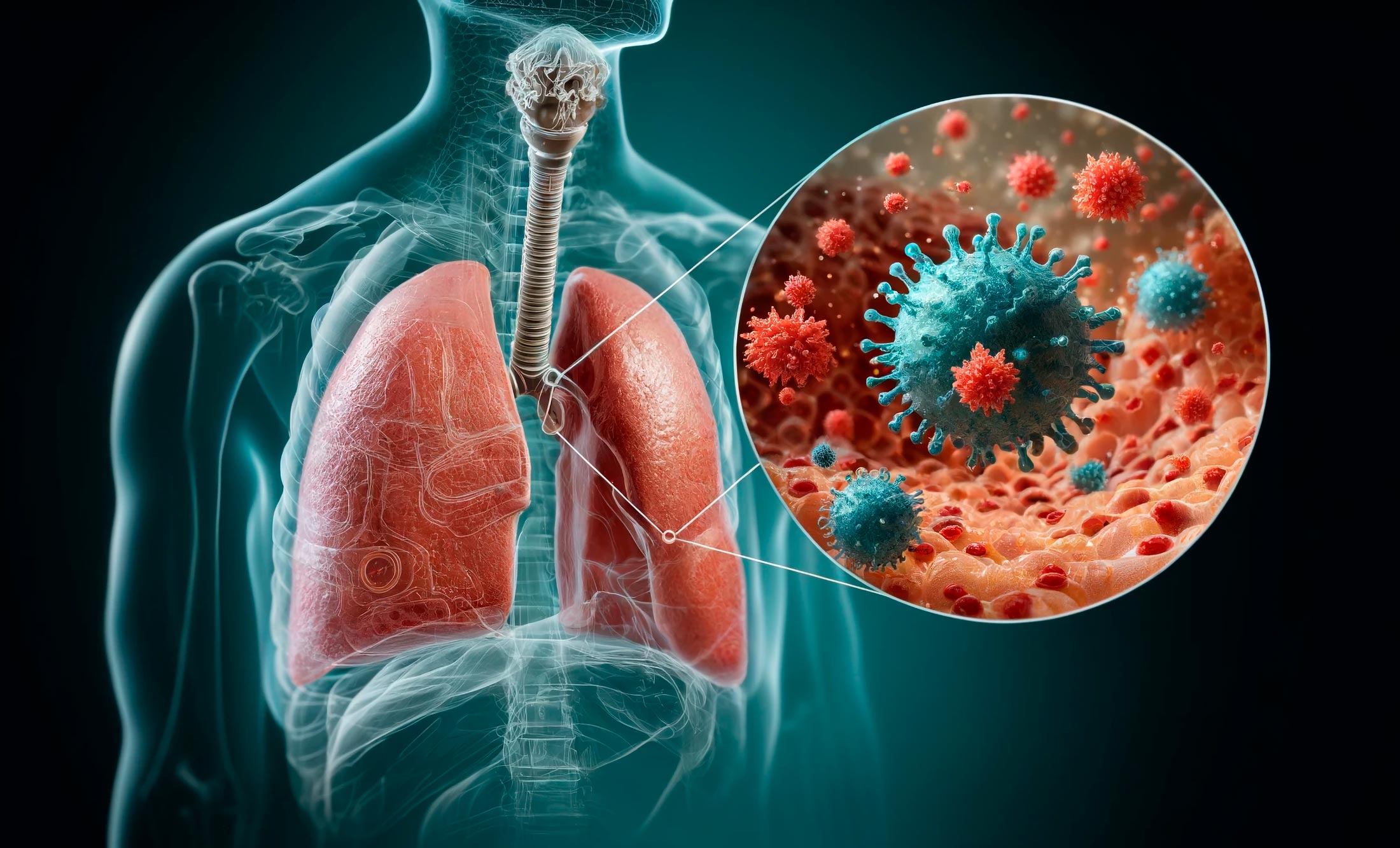
Stanford Medicine Unmasks the Surprising Instigators of Severe COVID in the Lungs
A previously overlooked type of immune cell allows SARS-CoV-2 to proliferate, Stanford Medicine scientists have found. The discovery has important implications for preventing severe COVID-19. The lung cell type that’s most susceptible to infection by SARS-CoV-2, the virus that causes COVID-19, is n
scitechdaily.com
Stanford Medicine Unmasks the Surprising Instigators of Severe COVID in the Lung
By Bruce Goldman, Stanford Medicine
April 16, 2024
Research shows interstitial macrophages are unexpectedly the primary target for SARS-CoV-2 in the lungs, leading to severe COVID-19, suggesting new therapeutic targets. Credit: SciTechDaily.com
A previously overlooked type of immune cell allows SARS-CoV-2 to proliferate, Stanford Medicine scientists have found. The discovery has important implications for preventing severe COVID-19.
The lung cell type that’s most susceptible to infection by SARS-CoV-2, the virus that causes COVID-19, is not the one previously assumed to be most vulnerable. What’s more, the virus enters this susceptible cell via an unexpected route. The medical consequences may be significant.
Stanford Medicine investigators have implicated a type of immune cell known as an interstitial macrophage in the critical transition from a merely bothersome COVID-19 case to a potentially deadly one. Interstitial macrophages are situated deep in the lungs, ordinarily protecting that precious organ by, among other things, engorging viruses, bacteria, fungi and dust particles that make their way down our airways. But it’s these very cells, the researchers have shown in a study that was published on April 10 in the Journal of Experimental Medicine, that of all known types of cells composing lung tissue are most susceptible to infection by SARS-CoV-2.
SARS-CoV-2-infected interstitial macrophages, the scientists have learned, morph into virus producers and squirt out inflammatory and scar-tissue-inducing chemical signals, potentially paving the road to pneumonia and damaging the lungs to the point where the virus, along with those potent secreted substances, can break out of the lungs and wreak havoc throughout the body.
The surprising findings point to new approaches in preventing a SARS-CoV-2 infection from becoming a life-threatening disease. Indeed, they may explain why monoclonal antibodies meant to combat severe COVID didn’t work well, if at all — and when they did work, it was only when they were administered early in the course of infection, when the virus was infecting cells in the upper airways leading to the lungs but hadn’t yet ensconced itself in lung tissue.

In an uninfected interstitial macrophage, the nucleus (purple) and outer cell membrane (blue) are intact. In an infected interstitial macrophage, the nucleus is shattered, copious newly made viral components (red) clump together, and the cell broadcasts inflammatory and scar-tissue-inducing chemical signals (yellow). Credit: Emily Moskal
The Virus Surprises
“We’ve overturned a number of false assumptions about how the virus actually replicates in the human lung,” said Catherine Blish, MD, PhD, a professor of infectious diseases and of microbiology and immunology and the George E. and Lucy Becker Professor in Medicine and associate dean for basic and translational research.Blish is the co-senior author of the study, along with Mark Krasnow, MD, PhD, the Paul and Mildred Berg Professor of biochemistry and the Executive Director of the Vera Moulton Wall Center for Pulmonary Vascular Disease.
“The critical step, we think, is when the virus infects interstitial macrophages, triggering a massive inflammatory reaction that can flood the lungs and spread infection and inflammation to other organs,” Krasnow said. Blocking that step, he said, could prove to be a major therapeutic advance. But there’s a plot twist: The virus has an unusual way of getting inside these cells — a route drug developers have not yet learned how to block effectively — necessitating a new focus on that alternative mechanism, he added.
In a paper published in Nature in early 2020, Krasnow and his colleagues including then-graduate student Kyle Travaglini, PhD — who is also one of the new study’s co-lead authors along with MD-PhD student Timothy Wu — described a technique they’d worked out for isolating fresh human lungs; dissociating the cells from one another; and characterizing them, one by one, on the basis of which genes within each cell were active and how much so. Using that technique, the Krasnow lab and collaborators were able to discern more than 50 distinct cell types, assembling an atlas of healthy lung cells.
“We’d just compiled this atlas when the COVID-19 pandemic hit,” Krasnow said. Soon afterward, he learned that Blish and Arjun Rustagi, MD, PhD, instructor of infectious diseases and another lead co-author of the study, were building an ultra-safe facility where they could safely grow SARS-CoV-2 and infect cells with it.
A collaboration ensued. Krasnow and Blish and their associates obtained fresh healthy lung tissue excised from seven surgical patients and five deceased lung donors whose lungs were virus-free but for one reason or another not used in transplants. After infecting the lung tissue with SARS-CoV-2 and waiting one to three days for the infection to spread, they separated and typed the cells to generate an infected-lung-cell atlas, analogous to the one Krasnow’s team had created with healthy lung cells. They saw most of the cell types that Krasnow’s team had identified in healthy lung tissue.
Now the scientists could compare pristine versus SARS-CoV-2-infected lungs cells of the same cell type and see how they differed: They wanted to know which cells the virus infected, how easily SARS-CoV-2 replicated in infected cells, and which genes the infected cells cranked up or dialed down compared with their healthy counterparts’ activity levels.
They were able to do this for each of the dozens of different cell types they’d identified in both healthy and infected lungs.
“It was a straightforward experiment, and the questions we were asking were obvious,” Krasnow said. “It was the answers we weren’t prepared for.”
It’s been assumed that the cells in the lungs that are most vulnerable to SARS-CoV-2 infection are those known as alveolar type 2 cells. That’s because the surfaces of these cells, along with those of numerous other cell types in the heart, gut and other organs, sport many copies of a molecule known as ACE2. SARS-CoV-2 has been shown to be able to grab onto ACE2 and manipulate it in a way that allows the virus to maneuver its way into cells.
Alveolar type 2 cells are somewhat vulnerable to SARS-CoV-2, the scientists found. But the cell types that were by far the most frequently infected turned out to be two varieties of a cell type called a macrophage.
Virus Factories
The word “macrophage” comes from two Greek terms meaning, roughly, “big eater.” This name is not unearned. The air we inhale carries not only oxygen but, unfortunately, tiny airborne dirt particles, fungal spores, bacteria and viruses. A macrophage earns its keep by, among other things, gobbling up these foreign bodies.The airways leading to our lungs culminate in myriad alveoli, minuscule one-cell-thick air sacs, which are abutted by abundant capillaries. This interface, called the interstitium, is where oxygen in the air we breathe enters the bloodstream and is then distributed to the rest of the body by the circulatory system.
The two kinds of SARS-CoV-2-susceptible lung-associated macrophages are positioned in two different places. So-called alveolar macrophages hang out in the air spaces within the alveoli. Once infected, these cells smolder, producing and dribbling out some viral progeny at a casual pace but more or less keeping a stiff upper lip and maintaining their normal function. This behavior may allow them to feed SARS-CoV-2’s progression by incubating and generating a steady supply of new viral particles that escape by stealth and penetrate the layer of cells enclosing the alveoli.
Interstitial macrophages, the other cell type revealed to be easily and profoundly infected by SARS-CoV-2, patrol the far side of the alveoli, where the rubber of oxygen meets the road of red blood cells. If an invading viral particle or other microbe manages to evade alveolar macrophages’ vigilance, infect and punch through the layer of cells enclosing the alveoli, jeopardizing not only the lungs but the rest of the body, interstitial macrophages are ready to jump in and protect the neighborhood.
At least, usually. But when an interstitial macrophage meets SARS-CoV-2, it’s a different story. Rather than get eaten by the omnivorous immune cell, the virus infects it.
And an infected interstitial macrophage doesn’t just smolder; it catches on fire. All hell breaks loose as the virus literally seizes the controls and takes over, hijacking a cell’s protein- and nucleic-acid-making machinery. In the course of producing massive numbers of copies of itself, SARS-CoV-2 destroys the boundaries separating the cell nucleus from the rest of the cell like a spatula shattering and scattering the yolk of a raw egg. The viral progeny exit the spent macrophage and move on to infect other cells.
But that’s not all. In contrast to alveolar macrophages, infected interstitial macrophages pump out substances that signal other immune cells elsewhere in the body to head for the lungs. In a patient, Krasnow suggested, this would trigger an inflammatory influx of such cells. As the lungs fill with cells and fluid that comes with them, oxygen exchange becomes impossible. The barrier maintaining alveolar integrity grows progressively damaged. Leakage of infected fluids from damaged alveoli propels viral progeny into the bloodstream, blasting the infection and inflammation to distant organs.
Yet other substances released by SARS-CoV-2-infected interstitial macrophages stimulate the production of fibrous material in connective tissue, resulting in scarring of the lungs. In a living patient, the replacement of oxygen-permeable cells with scar tissue would further render the lungs incapable of executing oxygen exchange.
“We can’t say that a lung cell sitting in a dish is going to get COVID,” Blish said. “But we suspect this may be the point where, in an actual patient, the infection transitions from manageable to severe.”
Another Point of Entry
Compounding this unexpected finding is the discovery that SARS-CoV-2 uses a different route to infect interstitial macrophages than the one it uses to infect the other types.Unlike alveolar type 2 cells and alveolar macrophages, to which the virus gains access by clinging to ACE2 on their surfaces, SARS-CoV-2 breaks into interstitial macrophages using a different receptor these cells display. In the study, blocking SARS-CoV-2’s binding to ACE2 protected the former cells but failed to dent the latter cells’ susceptibility to SARS-CoV-2 infection.
“SARS-CoV2 was not using ACE2 to get into interstitial macrophages,” Krasnow said. “It enters via another receptor called CD209.”
That would seem to explain why monoclonal antibodies developed specifically to block SARS-CoV-2/ACE2 interaction failed to mitigate or prevent severe COVID-19 cases.
It’s time to find a whole new set of drugs that can impede SARS-CoV-2/CD209 binding. Now, Krasnow said.
Reference: “Interstitial macrophages are a focus of viral takeover and inflammation in COVID-19 initiation in human lung ” by Timothy Ting-Hsuan Wu, Kyle J. Travaglini, Arjun Rustagi, Duo Xu, Yue Zhang, Leonid Andronov, SoRi Jang, Astrid Gillich, Roozbeh Dehghannasiri, Giovanny J. Martínez-Colón, Aimee Beck, Daniel Dan Liu, Aaron J. Wilk, Maurizio Morri, Winston L. Trope, Rob Bierman, Irving L. Weissman, Joseph B. Shrager, Stephen R. Quake, Christin S. Kuo, Julia Salzman, W.E. Moerner, Peter S. Kim, Catherine A. Blish and Mark A. Krasnow, 10 April 2024, Journal of Experimental Medicine.
DOI: 10.1084/jem.20232192
The study was funded by the National Institutes of Health (grants K08AI163369, T32AI007502, and T32DK007217), the Bill & Melinda Gates Foundation, Chan Zuckerberg Biohub, the Burroughs Wellcome Fund, Stanford Chem-H, the Stanford Innovative Medicine Accelerator, and the Howard Hughes Medical Institute.
Heliobas Disciple
TB Fanatic
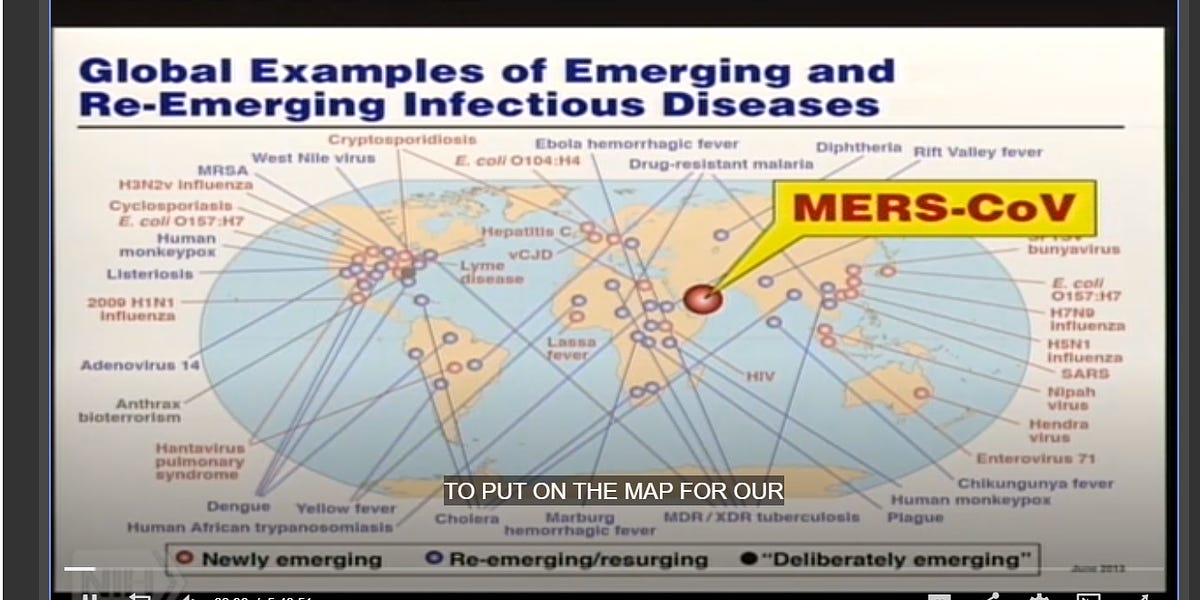
Why did Tony Fauci say under oath he barely knew top coronavirus scientist Ralph Baric - when in fact Fauci hosted a daylong 2013 meeting where Baric laid out his plans for risky research?
Peter Daszak - another scientist connected to China whom Fauci claimed under oath not to remember - was also at the 2013 meeting. Poor Tony, stuck with such a severe case of the "I don't recalls."
 alexberenson.substack.com
alexberenson.substack.com
Why did Tony Fauci say under oath he barely knew top coronavirus scientist Ralph Baric - when in fact Fauci hosted a daylong 2013 meeting where Baric laid out his plans for risky research?
Peter Daszak - another scientist connected to China whom Fauci claimed under oath not to remember - was also at the 2013 meeting. Poor Tony, stuck with such a severe case of the "I don't recalls."
Alex Berenson
Apr 12, 2024
(PART 1)
The evidence has been hiding in plain sight for years.
On June 24, 2013, Dr. Anthony S. Fauci invited a small group of top coronavirus researchers to an all-day brainstorming session at a conference center at the National Institutes of Health.
The subject was nominally an outbreak of a new illness called MERS, or Middle Eastern Respiratory Syndrome, a lung disease caused by a novel coronavirus. At the time, about 70 people had contracted the virus. Almost 40 had died, an eye-popping fatality rate, making MERS more lethal than smallpox.
Despite its lethality, MERS appeared to be a minor health threat, because it did not spread rapidly or easily. So the scientists - and one scientist in particular, Dr. Ralph S. Baric, the world’s top expert on coronaviruses - spent the day talking about threats from not just MERS and its potential evolution, but novel coronaviruses generally.
Fortunately, the NIH has preserved a complete video record of the conference, which lasted almost six hours (not including a lunch break). It is available here.
And, on many different levels, from beginning to end, it is astonishing.
—
The video directly refutes Fauci’s denial under oath of knowing Baric and Peter Daszak, the British zoologist who along with Baric was a key link between the American government and the Chinese virologists in Wuhan whose work likely caused the SARS-Cov-2 epidemic.
In a deposition in 2022, Fauci said of Baric, a North Carolina microbiologist who has studied coronaviruses for decades, “I doubt if I've ever met him.” He characterized Daszak as not even an “acquaintance.”
Yet Daszak and other officials in his group EcoHealth Alliance are present at the meeting, which Fauci opened with a speech. Nine months later, Fauci would again cross paths with Daszak, when Fauci spoke on a panel in Washington about emerging infectious diseases, part of a two-day conference that Daszak had moderated and helped create.
But as the world’s preeminent coronavirus researcher, Baric was even more important to Fauci. As he opened the June 2013 meeting, Fauci spoke openly of wanting to be sure he constantly had new viral threats that would ensure his research budget would continue to grow.
Nothing more perfectly illustrated the potential threat of emerging infectious disease than the coronavirus - a seemingly minor pathogen that suddenly had erupted into a lethal outbreak, the second in a decade.
“Now we have a new one [disease] to put on the map for our friends [in Congress] who support us in these rather constrained times,” Fauci said in his opening address. He wasn’t joking or speaking metaphorically - he showed the scientists the actual map.
—
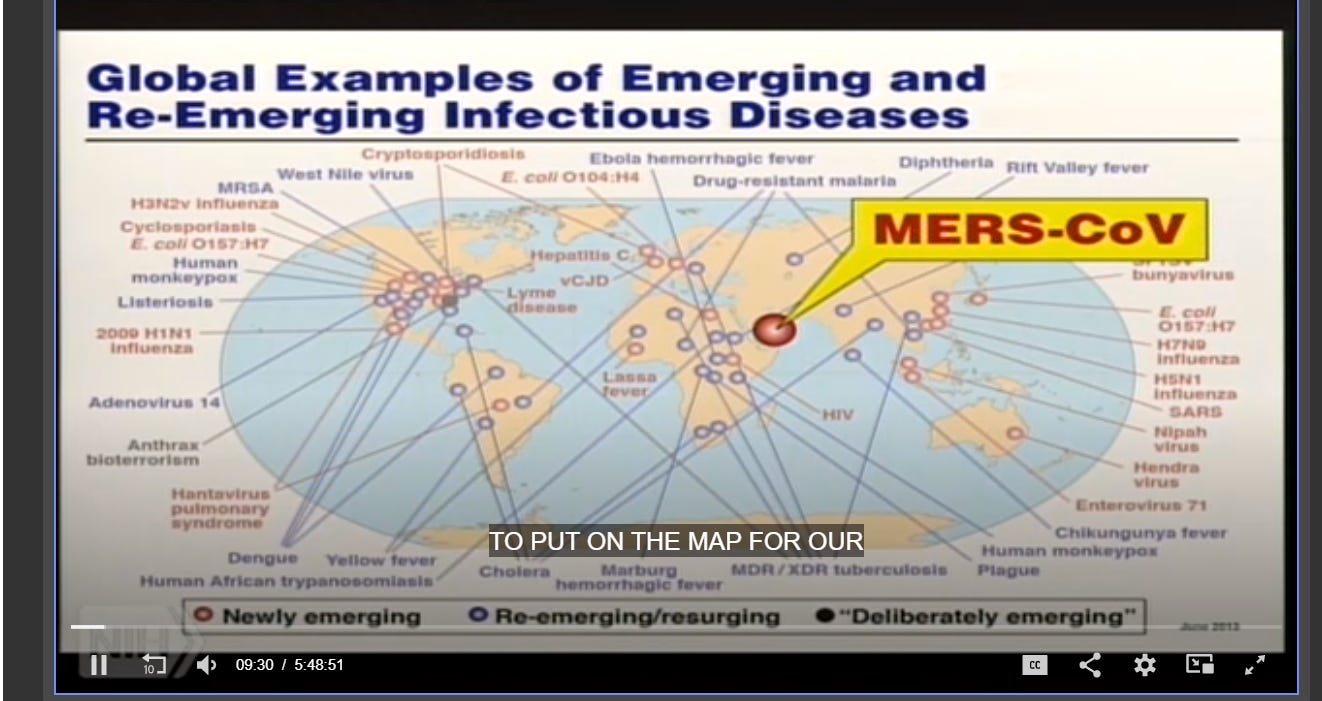
SOURCE
—
After his talk, Fauci took a seat alone in the front row to listen to the conference’s first invited speaker explain the growing threat of coronaviruses and how scientists and the NIH could work to fight them.
That speaker?
None other than Baric, who went on to dominate the day’s discussion. Not only did he give the first presentation, he returned after lunch to give another. He took over questioning repeatedly. One of the other scientists referred to him as an “eminence grise.” Another joked the NIH would be cutting Baric a check for a million dollars, no strings attached.
No wonder that seven years later, in February 2020, as Covid exploded out of China, Fauci would spend his extremely valuable time bring Baric into his offices for another private meeting to discuss the new epidemic - and its possible origins. Under oath, Fauci claimed not to remember anything about that meeting either, though it had taken place barely two years before.
—
(From page 32 of Dr. Anthony S. Fauci’s deposition in Missouri v Biden, given under oath on Nov. 23, 2022.)

SOURCE
—
So just why would Fauci go so far out of his way to deny knowing Baric? Why make such an implausible claim?
The rest of the video from June 24, 2013 offers as good an explanation as any, showing just how far Baric was willing go in trying to bring coronaviruses to heel - even if doing so raised the risk of a lab-created epidemic far deadlier than one that any natural coronavirus had ever caused.
(END OF PART 1)
Heliobas Disciple
TB Fanatic

Ten years ago, microbiologist Ralph Baric told Tony Fauci and the world he would make coronaviruses more lethal. Then - with Fauci's backing - he did.
Baric, who worked hand-in-glove with the lab in Wuhan from which Sars-Cov-2 likely leaked, also insisted the government keep its rules on coronaviruses loose so he could tinker with them more easily.
alexberenson.substack.com
Ten years ago, microbiologist Ralph Baric told Tony Fauci and the world he would make coronaviruses more lethal. Then - with Fauci's backing - he did.
Baric, who worked hand-in-glove with the lab in Wuhan from which Sars-Cov-2 likely leaked, also insisted the government keep its rules on coronaviruses loose so he could tinker with them more easily.
Alex Berenson
Apr 15, 2024
(SECOND OF TWO PARTS; PART ONE IS HERE)
Questioned under oath in 2022, Dr. Anthony S. Fauci largely denied knowing Dr. Ralph S. Baric, a North Carolina scientist and the world’s top coronavirus researcher.
Fauci’s claim came though he met one-on-one with Baric as Covid raced around the globe in early 2020 - and though he had funded Baric’s work and hosted Baric at a day-long coronavirus research conference in 2013. (I wrote about Fauci’s unlikely memory lapses here, in Part One of this piece.)
Why did Fauci try so hard to insulate himself from Baric 18 months ago - even at the cost of giving nonsensical answers while under oath?
—
Baric’s role and the plans he outlined in the conference on June 24, 2013 may help answer that question.
As I explained in Part One, the National Institutes of Health put the day-long event together during an outbreak of a novel coronavirus called MERS-Cov, which was highly lethal but not very transmissible.
A complete video of the conference is publicly available here. For anyone interested in understand the science - and the scientists - that may have led to Covid, it is a six-hour must-watch.
About 50 people attended the event. They included leading coronavirus researchers, as well as Peter Daszak and members of his EcoHealth Alliance, and federal officials concerned about infectious diseases. But Baric was without doubt the star of the show.
He made the first presentation, then another after lunch. He rarely went more than a few minutes without asking questions. He had earned the role. After earning a PhD in microbiology from North Carolina State University in 1982, Baric turned to infectious disease research.
He became known for his ability to handle coronaviruses, which can be hard to grow in labs, as well as in making genetic modifications to them. Because coronaviruses have a relatively large genome - the nucleotides that encode the information necessary for them to replicate themselves - they are harder to genetically modify than some other viruses. But Baric and his researchers did so with relative ease.
—
(Destroying the world in order to save it, a presentation by Dr. Ralph S. Baric. Yes, that’s Tony Fauci in the front row.)

SOURCE
—
Coronaviruses are named for their striking shape. They have a halo of “spike” proteins sticking out of a shell that protects the strand of genes that lets them replicate. The spikes attach to receptors on human cells, letting the coronavirus begin the process of copying itself.
Doctors had historically considered coronaviruses minor health threats compared to influenza, much less terrors like smallpox. The four coronaviruses known to infect humans before 2002 were generally associated with common colds.
Most respiratory virus researchers focused on influenza, whose dangers were more obvious. But the 2002-03 SARS epidemic, which killed about 800 people and caused a worldwide panic, and the MERS outbreak a decade later, made coronaviruses a hot topic among infectious disease researchers - and the policymakers who funded them.
Baric made sure everyone understood the risk of his favorite virus - and the need for well-funded research programs to reduce it.
Sure, neither SARS nor MERS appeared very transmissible, with MERS in particular seemingly requiring prolonged and close contact.
But their fatality rates were striking — 10 percent for SARS, 50 percent for MERS. With a few genetic modifications, similar coronaviruses might spread far more quickly, Baric argued.
—
The best way to predict and ultimately reduce that risk would be to build coronaviruses more lethal and transmissible than any yet found in nature, Baric said in response to a question near the end of his first presentation:
The other way to do it… [is] make in essence recombinants where you drop the spike gene in different bat coronaviruses which you think may have appropriate properties that could be emergent, preemergent strains and characterize their ability to replicate in human cells…
Baric then acknowledged such work might be so risky that it might fall into the category of “dual-use experiments,” meaning research that militaries or terrorists might use to create deadlier biological weapons.
“Now, maybe dual-use experiments of concern, that’s another issue,” he said, with a slight giggle. (Yes, the video makes clear he does giggle, at approximately 45:55.)
—
(Do-it-yerself coronaviruses, the Ralph Baric way! From a 2006 Baric paper titled “Synthetic Viral Genomics: Risks and Benefits for Science and Society.”)
—
At a 2014 conference in Washington, D.C., Baric would also acknowledge his laboratory work outran any natural mutations that coronaviruses so far discovered in living animals had managed to produce.
After discussing three potential ways for the viruses to broaden their “host range” — that is, infect different species of animals, including humans — Baric acknowledged that two of the three “have only been identified using in vitro models, although the potential exists for similar mutants to emerge naturally in nature.”
Why do this work at all?
Baric and other scientists at the 2013 conference explained their work as necessary both to understand the potential threat from coronaviruses and to defuse it by helping research on antiviral drugs and vaccines.
—
Never mind that the despite the publicity the SARS and MERS outbreaks had garnered, theor combined global death toll equaled roughly one day of flu deaths.
Never mind that vaccines for flu, the closest counterpart to coronaviruses, appeared mostly useless and that decades of research had not brought the world closer to better ones.
By making stickier spikes and fitter, more dangerous viruses, Baric would enable the development of better vaccines and antivirals. He would create an arms race, in order to win it.
Baric never put it quite this bluntly, but near the end of the conference - five hours and 26 minutes in, according to the video - he came close. “It’s basically structure-based antigen design,” he said. “You can blend S [spike] components… to provide a broader neutralizing response within that cluster.”
—
Baric wasn’t just outlining potential research pathways in June 2013. He intended to do the work, and he did. In November 2015, he — along with a Chinese virologist named Shi Zhengli — published what became a notorious paper in Nature Medicine:
A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence
Baric, Shi, and the other authors explained they had “generated” a “virus expressing the spike of bat coronavirus SHC014 in a mouse-adapted SARS-CoV backbone.”In other words, Baric had combined multiple coronaviruses to make one better at infecting humans. The new lab-made virus would “replicate efficiently in primary human airway cells,” the paper explained. “Additionally, in vivo experiments demonstrate replication of the chimeric virus in mouse lung with notable pathogenesis.”
Baric’s co-author Shi was, of course, none other than the head of coronavirus research at the Wuhan Institutes of Virology, the lab from which Covid likely leaked in 2019. In 2018, Daszak’s EcoHealth Alliance attempted to get federal funding for a new project where the Wuhan lab and Baric would again collaborate on work to make coronaviruses more dangerous.
The United States government rejected the request as too risky; but whether Shi’s group went ahead with the research anyway and accidentally created Sars-Cov-2 is the most important question about Covid’s origins. The second most important is what Baric, Daszak, and other American researchers knew or suspected about Shi’s research as Covid exploded worldwide.
(In 2020, Nature Medicine added a defensive, if snippy, note to the article complaining that it was “being used as the basis for unverified theories that the novel coronavirus causing COVID-19 was engineered. There is no evidence that this is true.”)
—
In 2016, Baric was the senior author on a paper in the Proceedings of the National Academy of Sciences that arguably went even further than the Nature Medicine paper. The research group he led had manipulated a coronavirus provided by Shi and found it “poised for human emergence.”
The authors explained that they had “construct[ed] chimeric and full-length zoonotic coronaviruses to evaluate emergence potential… the results indicate a significant threat posed by WIV1-CoV.” The lab-grown viruses “replicated efficiently in human airway cultures,” they wrote.
—
Within the biomedical research community, research like Baric’s was controversial even at the time.
In a 2011 paper, virologists had shown that they could make avian influenza transmissible between ferrets, who are considered excellent animal models for humans. Avian flu does not easily infect humans, but it can be highly lethal when it does, so the avian flu experiment stirred serious concerns.
But the infectious disease researchers had an important ally: Tony Fauci, who wrote in the Washington Post in 2011 that the flu research was a “risk worth taking.”
Still, the obvious risks posed by highly infectious and dangerous viruses like smallpox had led the federal government to create what it called a list of select agents which required special precautions. Besides smallpox, the list included anthrax, plague, and the original SARS.
But MERS, despite its lethality, was not considered a “select agent”. And in the final moments of the June 24, 2013 conference, Ralph Baric called for the government not add it to the list.
“If you want fast movement forward, and you restrict it with select agent status, then that will have a very chilling effect on productivity,” Baric said. “I think somebody had to say it.”
MERS was never added to the select agent list.
And six-and-half-years later, the massive and deadly coronavirus epidemic that Baric had predicted - and worked so hard to stop - exploded around the globe.
(END OF PART TWO)
Heliobas Disciple
TB Fanatic

Whites Twice as Likely to Die of COVID in California Than Non-Whites
A mysterious finding
 www.igor-chudov.com
www.igor-chudov.com
Whites Twice as Likely to Die of COVID in California Than Non-Whites
A mysterious finding
Igor Chudov
Apr 16, 2024
San Jose Mercury News reports an unusual COVID pattern detected in California.

What is interesting is that despite comprising only 37% of Californians, White people account for 60% of all COVID deaths during the recent period.
The article compares Feb-Aug 2020 (highlighted in blue) vs. Sep 2023 through Feb 2024 (highlighted in red).
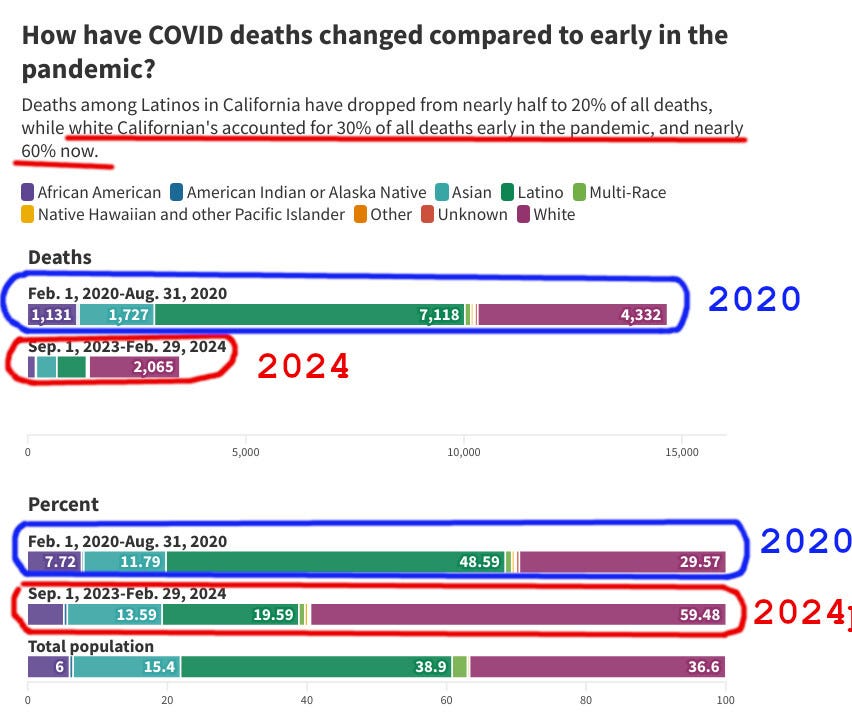
The results for 2020 confirm a modest advantage that we would expect from the “white privilege”: Whites comprised only 30% of deaths despite being 37% of the population.
Whites were able to mitigate the effects of the pandemic due to a more privileged position: belonging to the ”laptop class,” not working in as many customer-facing jobs, having better medical insurance, living in less dense housing, and so on. So, it is not a surprise that in 2020, White people were slightly less likely to die from Covid (30% of deaths despite being 37% of the population).
The real surprise is what is happening in 2024. While the overall number of COVID deaths thankfully declined, deaths among White people (59.48%) account for much more than their share of the population (37%)!
The Whites’ share of COVID deaths doubled from 30% in 2020 to 60% in 2024.
Why would that be?
COVID Vaccines
Is it a coincidence that White people are also more vaccinated and boosted against COVID-19?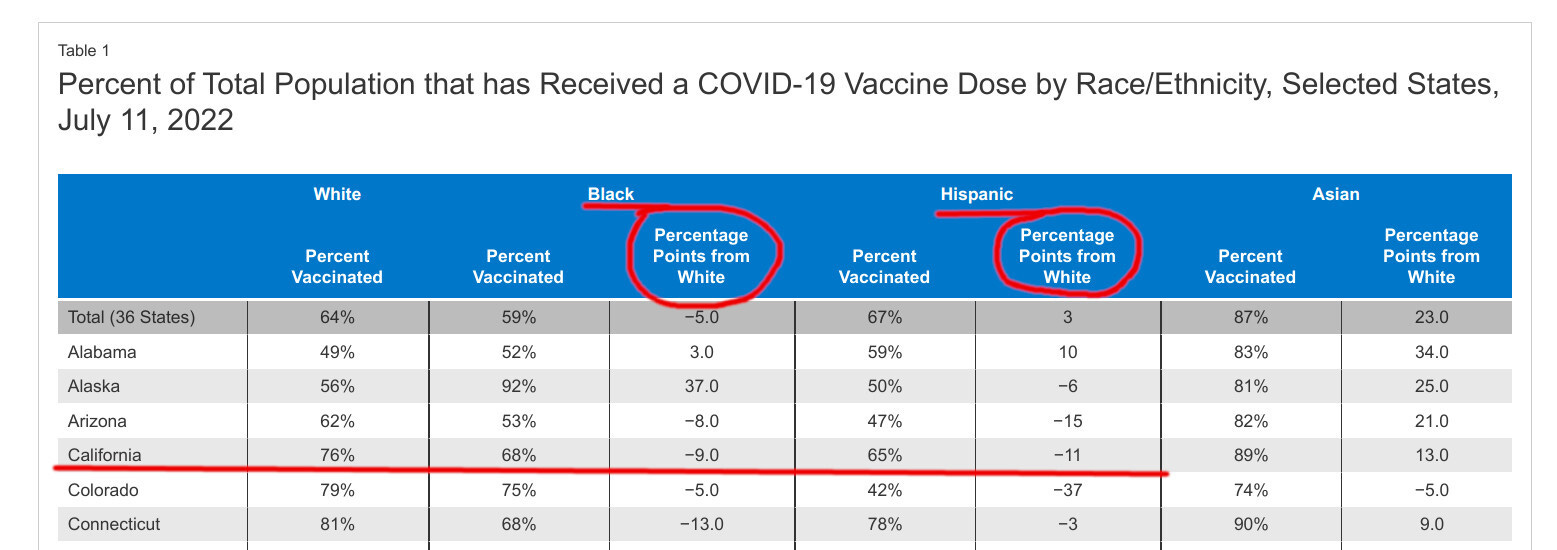
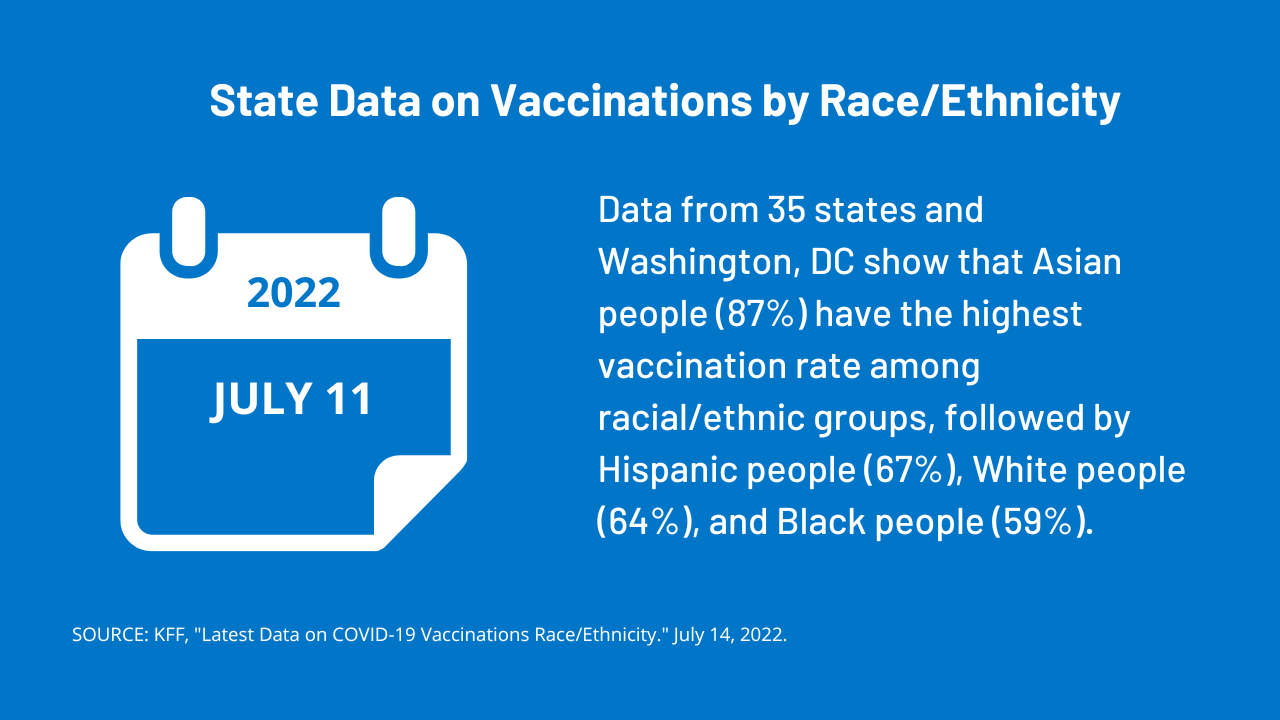
Latest Data on COVID-19 Vaccinations by Race/Ethnicity | KFF
KFF is collecting and analyzing data on COVID-19 vaccinations by race/ethnicity to gain increased insight who is receiving the vaccine and whether some groups are facing disparities in vaccination.
 www.kff.org
www.kff.org
The more vaccinated and boosted race in California is accounting for GREATER share of Covid deaths compared to their share of the population. Meanwhile, Black and Hispanic Californians, who are less vaccinated, account for fewer deaths than their share of the population would suggest.
A coincidence? If so, it continues a long line of other subpopulations where higher vaccination rates are associated with greater mortality:
Igor’s Newsletter
Excess Mortality Still Correlated With COVID Vaccination Rate - Dec 2023 data
December 2023 was unusually but not unexpectedly severe: a new COVID variant, JN.1, swept Europe and the rest of the world. Other pathogens circulated as well, in addition to the usual killers such as cancer, heart disease, blood clots, dementia, and so on. As a result, Europe experienced a month of…
Read more
a month ago · 426 likes · 297 comments · Igor Chudov
Remember the economic advantages I mentioned that made White people have fewer deaths in 2020? Those have not changed in 2024: the laptop class, better medical care, less dense housing, etc, are still disproportionally available to White people.
And yet, despite those advantages, in 2024, Whites are more likely to die of Covid than their less privileged, and also less vaccinated, Hispanic and Black counterparts! The “white privilege” mysteriously stopped working after the whites were disproportionately vaccinated.
We were told that Covid vaccines save lives. If so, can we ask why they make the groups that received more of them more likely to suffer COVID deaths?
Any thoughts?
Heliobas Disciple
TB Fanatic

How will the ongoing immune escape pandemic end?
Large-scale pharmaceutical and non-pharmaceutical interventions in a natural pandemic of an acute, self-limiting viral infection drive the dominant propagation of more infectious immune escape variants. This complicates and delays Nature’s control over the pandemic. Once the train of immune...
 voiceforscienceandsolidarity.substack.com
voiceforscienceandsolidarity.substack.com
How will the ongoing immune escape pandemic end?
Geert Vanden Bossche
Apr 15, 2024
Large-scale pharmaceutical and non-pharmaceutical interventions in a natural pandemic of an acute, self-limiting viral infection drive the dominant propagation of more infectious immune escape variants. This complicates and delays Nature’s control over the pandemic. Once the train of immune escape gains momentum, highly Covid-19 (C-19) vaccinated populations will inevitably reach the terminus of high virulence, referred to as 'HIVICRON'. Consequently, the toll exacted by this interference on the population by Nature will be MUCH higher compared to if human intervention had been limited to appropriately treating vulnerable individuals at an early stage of symptomatic infection. The losses incurred by large-scale pharmaceutical and non-pharmaceutical interventions are attributable to the immune evasion they induce. This indirectly leads to the derailment of the immune response, while simultaneously causing a surge in breakthrough infections that further extend the immune evasion to more conserved domains within the spike protein and other viral proteins in general. This causes highly C-19 vaccinated populations to initially exert high immune pressure on the infectiousness of the virus, and ultimately on its overall transmission potential. This progression inevitably culminates in a final stage where the immune pressure on the virus's capacity to propagate and survive rises to an extent that it is compelled to spread massively within the host rather than from one host to another. I have no doubt that this evolution will result in a rapid dissemination of the virus from the upper respiratory tract to all organs, leading to swift and extensive mortality in these populations.
In conclusion, only after causing some painful detours and acquiring a high level of virulence in highly C-19 vaccinated populations, will population immunity eventually end the pandemic of immune escape and drive the virus into endemicity (as illustrated in fig. A appended below). This sharply contrasts with how a natural SARS-CoV-2 pandemic would have transitioned into endemicity (as illustrated in fig. B below)


Heliobas Disciple
TB Fanatic

The microcosm of those who crown themselves as emperors of science...(with translation in Dutch!)
The microcosm of those who crown themselves as emperors of science... Recently, a professor at the University of Antwerp criticized me; he questioned my scientific credibility because, according to him, I haven't published enough in peer-reviewed journals!
voiceforscienceandsolidarity.substack.com
The microcosm of those who crown themselves as emperors of science...
Geert Vanden Bossche
Apr 16, 2024
The microcosm of those who crown themselves as emperors of science...
Recently, a professor at the University of Antwerp criticized me; he questioned my scientific credibility because, according to him, I haven't published enough in peer-reviewed journals!
It just goes to show that many of our scientists and so-called health experts live in a microcosm where everything seems to revolve solely around publications. Despite their relentless efforts to continually plant new trees and explore those planted by others, they are losing sight of the forest itself and getting lost in it. This has been clearly exemplified by their misjudgment regarding the COVID-19 situation and the mass vaccination they blindly and unquestioningly supported.
Every year, several billions of dollars of taxpayer money are pumped into their research worldwide, without the taxpayer, or society, benefiting in any way. They constantly exercise in collecting puzzle pieces but fail to show any or enough intellectual diligence to piece them together to understand complex and multifaceted phenomena, such as the interaction between a virus and the immune response of a host population. Then, when important questions arise about whether interventions are meaningful or not, they suddenly want to venture into the macrocosm of science where they've never been before. Even though for many of them this subject matter is foreign and extends far beyond their knowledge silo, they still trust that they somehow have science in their pocket and that their 'knowledge' conveniently aligns with the financial interests of their sponsors. Hence, they believe it's completely unnecessary, even counterproductive, to engage in a debate with scientists who think differently, especially when these scientists don't excel in producing publications. What surprises me the most is that their shortsightedness and urge to lead the race at all costs reach so far that they are not even aware that their shortcuts in immunology and vaccinology not only miss efficiency but also put the entire peloton in great danger.
Heliobas Disciple
TB Fanatic
IF YOU ONLY HAVE TIME TO READ ONE POST I POSTED TODAY, READ THIS ONE.
You don't have to understand all the science (I surely don't), but keep reading past what you don't understand so you get the general picture. Radagast is reaching the same conclusion Geert reaches, although I'm not sure if their way to get to that conclusion is the same because the science is over my head and I don't have the patience to try to figure it out like I used to do with Geert (I don't try anymore with him either, I realized after hours listening and figuring out the science after a lot of concentration that it doesn't matter if I follow it all to get the gist of what they're saying). It's worth reading even if we don't understand all of the nitty gritty. I would love if Phillip McMillan would interview both of them together and get a discussion going! (and break it down so we can all understand all of it)
~~~~~~~~~~~~~~~~~
(fair use applies)
Hey look, serotypes!
Radagast
April 16, 2024
I have to point this out. About half a year ago, I said that you’re probably going to end up with SARS2 serotypes, simply because it was unlikely by now that something new that emerges would be superior enough to completely eradicate the XBB lineages. Immunity would first be reoriented against the new serotype and that would then create a landscape within which the old serotype could re-emerge.
Well guess what happened. It turns out there’s an XBB lineage with a weird insertion at 212 that emerged in Canada and spread around the globe. So now you have two versions of the virus, both with insertions in different parts of the NTD. They’re both going to be under pressure to change the NTD in different ways. For JN.1 it’s straightforward to start deleting stuff around the first loop, due to the insertion at 16. We see it happening in multiple JN.1 lineages, a deletion at 31, just after the first immunogenic loop and a bit after the insertion, that allows a new glycan to be added at 30. But this XBB variant would have more of an incentive to delete chunks further along in the NTD.
Remember, it’s when deletions and new glycans emerge in the NTD, that the last antibodies that neutralize the virus in most of the population are going to fail. You will end up with very fusogenic variants, that spread by moving from one cell into another, without ever being in reach of the antibodies. The antibody response against the RBD is already pretty much useless by now.
The important thing to keep in mind is that once you have multiple of these highly fusogenic serotypes circulating in the population, you can’t vaccinate anymore. Most of the population already has high concentrations of antibodies, vaccination under these conditions won’t improve the population’s protection, it will merely favor one serotype at the cost of the other. By now it’s clear that the BA.2.86* Spike doesn’t do a clean sweep: It stays stuck around 93%, so it doesn’t completely wipe out the other lineages.
There are probably other serotypes out there too, we just don’t see them because sequencing is down by 99%. There’s a weird one in France, but nobody knows how widespread it is. There’s also a derivative of the original Omicron still circulating in Nebraska, infecting multiple people.
I know you’ve probably seen it argued there will be some sudden new doomsday variant this summer that leads to overnight vaccine failure. I don’t think this is likely, although it is inherently impossible to rule out.
In the coming weeks we can expect S:R346T, S:F456L, S:S31del to grow globally dominant.
Right now, adding the whole thing together suggests a 117% growth advantage.
What seems far more likely to me unfortunately, is that we’re gradually going to see different serotypes emerge: XBB lineages and BA.2.86 lineages, along with lineages based on whatever else is still out there.
Those lineages will undergo deletions in the immunogenic loops of the N-Terminal Domain, facilitated by the rare insertions they tend to develop in the NTD. These deletions will increase fusogenicity and thereby increase the intrinsic virulence of the virus.
That ultimately results in a virus that behaves more like the original SARS.
There is a notable difference by now between vaccinated and unvaccinated people, in regards to the regions their T cell receptors look for. This has been studied, in people who were infected in the first half of 2022.
Among the unvaccinated, they found strong overrepresentation of the S673-699 epitope. On the other hand, among the vaccinated, they found strong overrepresentation of these four regions:
S135-177, S264-276, S319-350, and S448-472.
The first of these four is easy to understand: It’s the N3 loop of the NTD. The other three regions are also in the NTD, but don’t correspond neatly to any of the known immunogenic loops.
But here’s the whole picture, which tells the story in my opinion:

I would argue it’s pretty obvious what’s going on here. The unvaccinated adaptive immune response tends to focus on regions in S2. The vaccinated adaptive immune response tends to focus on the N-Terminal Domain. We also see it strongly preferentially focuses on the region around 180, which is an infection enhancing site. The N5 loop also seems to be preferred.
So it seems straightforward, to expect that the NTD is going to undergo radical changes now, now that antibodies against the RBD have become useless. As the loops in the NTD grow shorter, by deletion mutations, you end up with a virus that behaves more like SARS1.
The entire Spike protein is covered by glycans, with the exception of the receptor binding domain. But now that the RBD evades the antibodies, we’re reaching the point where the antibody response depends on those last few regions of the NTD not covered by glycans, where antibodies can still bind to neutralize viral particles.
You also have to keep in mind: As more and more of the Spike protein becomes inaccessible to the T-cells, whether due to evolution to resemble our own proteins, deletions, steric hindrance, or glycans that are added, you’re going to see people’s lungs populated by an increasingly homogeneous population of T-cells.
With every breakthrough infection, the immunological abnormalities further escalate. That’s why you now see a very different antibody response against JN.1, when comparing vaccinated and unvaccinated people, as the vaccinated keep recalling antibodies against recessive epitopes in the NTD. This is what leads to the current situation, in which people suffer constantly elevated levels of respiratory disease. A lung environment populated with T cells looking for SARS2 has little room for other T cells looking for unrelated respiratory pathogens.
Through vaccination, you put abnormal antibody pressure on the Spike protein. This abnormal antibody pressure leads to affinity escape (escaping low affinity antibodies by increasing ACE2 affinity). But now that the abnormal antibody pressure is so strongly concentrated on the NTD, you’re encouraging the immunogenic loops of the NTD to grow shorter, which is known to dramatically increase fusogenicity. In other words, you are selecting more virulent variants.
It’s very simple: You want to see a small number of high affinity antibodies, against select regions of the Spike protein, in people who suffered severe infections. Those antibodies will be against regions that are associated with virulence. As a result, people will gradually offer a selective advantage to variants of this virus that are less virulent. What you don’t want to see, is this yellow line in the whole population:

This is what happened to the four hCovs in our species: We had a pandemic long ago (including one in the 19th century), a bunch of people died, then it gradually grew less virulent. But here’s what’s happening to SARS2:

It has been gradually growing more virulent since the first Omicron, but we don’t really notice it yet, as the most vulnerable people have already died by now. This happens, because the population is unable to select against virulence, because there is an antibody response against regions of the Spike protein that would normally not be targeted.
Why?
Well, we can look at the attempt to vaccinate chickens against influenza in Asia. These vaccines began selecting new variants, they ended up breeding versions of influenza that are extremely deadly and focus on infecting the brain rather than the lungs. Those versions have now spread around the world and killed millions of wild birds, entire species have had more than half their population die in a single season.
SARS2 is happily proceeding along the same path, with vaccine resistant versions emerging that are getting better at infecting the brain and increasingly competent at fusing cells together. In both cases we’re dealing with a virus that has an abnormal polybasic cleavage site, allowing it to infect endothelial cells.
If we’ve tried the exact same experiment of vaccinating against rapidly mutating respiratory viruses in another species and ended up breeding deadly viruses that decimate wild bird populations, wouldn’t you expect that performing the same experiment in our own species also results in deadly viruses that decimate our population?
You might argue to me that people now have immunity against proteins other than Spike too. But I don’t think it’s going to be very relevant, because you can only neutralize viral particles through Spike.
But there’s one element I’ve never addressed: interfering defective viral particles. This is not difficult to understand. When a cell is infected with the virus and does not get killed, but tries to get rid of the virus on its own, it will contain defective viral proteins or viral RNA. When it then gets infected again later on with a new variant, it will produce defective viral particles, that stimulate the immune system without being able to complete the viral cycle. These defective particles interfere in the normal particles replicating.
NK cells are aggressive upon a first infection, destroying infected cells. Upon subsequent infections however, they tend to prefer telling infected cells to solve the problem themselves. They have receptors they can use, to detect large chunks of weird RNA, which they will then delete. This means you end up with some persistent broken RNA in those cells, only activated again upon reinfection.
That’s a normal part of how immunity is supposed to work. It has a small protective effect on unrelated viruses, but a major protective effect on closely related viruses. Once you vaccinate people, you tend to break this immune mechanism, as the cells capable of producing defective viral particles will get killed.
Once you make the antibodies and T cells do the job of dealing with this virus, you set the immune system up for failure, relying entirely on these two mechanisms that are very vulnerable to the effects of mutations to the Spike protein. Such failure could come very suddenly and unpredictably, as changes to cysteine bridges can cause the whole NTD to fold differently, thereby evading most antibodies.
So all these studies, arguing that the virus never leaves your body, are not discovering something shocking. They’re just discovering how the immune system works: It encourages infected cells to store broken genetic material of viruses, that interferes with their replication next time the virus shows up.
Sometimes this means the virus fails to spread in your own body, sometimes it means you pass on versions of a virus with reduced fitness. Vaccinate everyone with inactivated vaccines and you break this mechanism.
Adults with two shots of mRNA:

Unvaccinated adults:

The immune system is still stuck with an abnormally elevated antibody response to this virus. As a result, the virus continues to be under pressure to shorten its immunogenic NTD loops and increase its fusogenicity.
In contrast, the innate immune system will have to do the job among unvaccinated adults. These are mechanisms that select against virulence.
The situation we’re dealing with, is one where the intrinsic virulence of this virus will continue to rise, until the antibody pressure exerted by the vaccinated is diminished.
Everything I’m saying here is a natural outgrowth of how the immune system works.
The main purpose of antibodies is to exert selection against virulence associated epitopes. That’s why survivors of the 1918 influenza pandemic have antibodies that react with the 1918 pandemic, but not with modern influenza strains. These people who built up natural immunity, with antibodies against select virulence-associated epitopes, are the reason the 1918 strain of influenza was replaced over time by more benign strains: Their immune systems would only pass on more benign versions of influenza.
We broke that mechanism for our own short-term self-interest and made a handful of people billionaires in the process.
You don't have to understand all the science (I surely don't), but keep reading past what you don't understand so you get the general picture. Radagast is reaching the same conclusion Geert reaches, although I'm not sure if their way to get to that conclusion is the same because the science is over my head and I don't have the patience to try to figure it out like I used to do with Geert (I don't try anymore with him either, I realized after hours listening and figuring out the science after a lot of concentration that it doesn't matter if I follow it all to get the gist of what they're saying). It's worth reading even if we don't understand all of the nitty gritty. I would love if Phillip McMillan would interview both of them together and get a discussion going! (and break it down so we can all understand all of it)
~~~~~~~~~~~~~~~~~
(fair use applies)
Hey look, serotypes!
Radagast
April 16, 2024
I have to point this out. About half a year ago, I said that you’re probably going to end up with SARS2 serotypes, simply because it was unlikely by now that something new that emerges would be superior enough to completely eradicate the XBB lineages. Immunity would first be reoriented against the new serotype and that would then create a landscape within which the old serotype could re-emerge.
Well guess what happened. It turns out there’s an XBB lineage with a weird insertion at 212 that emerged in Canada and spread around the globe. So now you have two versions of the virus, both with insertions in different parts of the NTD. They’re both going to be under pressure to change the NTD in different ways. For JN.1 it’s straightforward to start deleting stuff around the first loop, due to the insertion at 16. We see it happening in multiple JN.1 lineages, a deletion at 31, just after the first immunogenic loop and a bit after the insertion, that allows a new glycan to be added at 30. But this XBB variant would have more of an incentive to delete chunks further along in the NTD.
Remember, it’s when deletions and new glycans emerge in the NTD, that the last antibodies that neutralize the virus in most of the population are going to fail. You will end up with very fusogenic variants, that spread by moving from one cell into another, without ever being in reach of the antibodies. The antibody response against the RBD is already pretty much useless by now.
The important thing to keep in mind is that once you have multiple of these highly fusogenic serotypes circulating in the population, you can’t vaccinate anymore. Most of the population already has high concentrations of antibodies, vaccination under these conditions won’t improve the population’s protection, it will merely favor one serotype at the cost of the other. By now it’s clear that the BA.2.86* Spike doesn’t do a clean sweep: It stays stuck around 93%, so it doesn’t completely wipe out the other lineages.
There are probably other serotypes out there too, we just don’t see them because sequencing is down by 99%. There’s a weird one in France, but nobody knows how widespread it is. There’s also a derivative of the original Omicron still circulating in Nebraska, infecting multiple people.
I know you’ve probably seen it argued there will be some sudden new doomsday variant this summer that leads to overnight vaccine failure. I don’t think this is likely, although it is inherently impossible to rule out.
In the coming weeks we can expect S:R346T, S:F456L, S:S31del to grow globally dominant.
Right now, adding the whole thing together suggests a 117% growth advantage.
What seems far more likely to me unfortunately, is that we’re gradually going to see different serotypes emerge: XBB lineages and BA.2.86 lineages, along with lineages based on whatever else is still out there.
Those lineages will undergo deletions in the immunogenic loops of the N-Terminal Domain, facilitated by the rare insertions they tend to develop in the NTD. These deletions will increase fusogenicity and thereby increase the intrinsic virulence of the virus.
That ultimately results in a virus that behaves more like the original SARS.
The reason this is happening
So why did this happen? Well, allow me to show you something.There is a notable difference by now between vaccinated and unvaccinated people, in regards to the regions their T cell receptors look for. This has been studied, in people who were infected in the first half of 2022.
Among the unvaccinated, they found strong overrepresentation of the S673-699 epitope. On the other hand, among the vaccinated, they found strong overrepresentation of these four regions:
S135-177, S264-276, S319-350, and S448-472.
The first of these four is easy to understand: It’s the N3 loop of the NTD. The other three regions are also in the NTD, but don’t correspond neatly to any of the known immunogenic loops.
But here’s the whole picture, which tells the story in my opinion:
I would argue it’s pretty obvious what’s going on here. The unvaccinated adaptive immune response tends to focus on regions in S2. The vaccinated adaptive immune response tends to focus on the N-Terminal Domain. We also see it strongly preferentially focuses on the region around 180, which is an infection enhancing site. The N5 loop also seems to be preferred.
So it seems straightforward, to expect that the NTD is going to undergo radical changes now, now that antibodies against the RBD have become useless. As the loops in the NTD grow shorter, by deletion mutations, you end up with a virus that behaves more like SARS1.
The entire Spike protein is covered by glycans, with the exception of the receptor binding domain. But now that the RBD evades the antibodies, we’re reaching the point where the antibody response depends on those last few regions of the NTD not covered by glycans, where antibodies can still bind to neutralize viral particles.
You also have to keep in mind: As more and more of the Spike protein becomes inaccessible to the T-cells, whether due to evolution to resemble our own proteins, deletions, steric hindrance, or glycans that are added, you’re going to see people’s lungs populated by an increasingly homogeneous population of T-cells.
With every breakthrough infection, the immunological abnormalities further escalate. That’s why you now see a very different antibody response against JN.1, when comparing vaccinated and unvaccinated people, as the vaccinated keep recalling antibodies against recessive epitopes in the NTD. This is what leads to the current situation, in which people suffer constantly elevated levels of respiratory disease. A lung environment populated with T cells looking for SARS2 has little room for other T cells looking for unrelated respiratory pathogens.
Through vaccination, you put abnormal antibody pressure on the Spike protein. This abnormal antibody pressure leads to affinity escape (escaping low affinity antibodies by increasing ACE2 affinity). But now that the abnormal antibody pressure is so strongly concentrated on the NTD, you’re encouraging the immunogenic loops of the NTD to grow shorter, which is known to dramatically increase fusogenicity. In other words, you are selecting more virulent variants.
It’s very simple: You want to see a small number of high affinity antibodies, against select regions of the Spike protein, in people who suffered severe infections. Those antibodies will be against regions that are associated with virulence. As a result, people will gradually offer a selective advantage to variants of this virus that are less virulent. What you don’t want to see, is this yellow line in the whole population:
This is what happened to the four hCovs in our species: We had a pandemic long ago (including one in the 19th century), a bunch of people died, then it gradually grew less virulent. But here’s what’s happening to SARS2:
It has been gradually growing more virulent since the first Omicron, but we don’t really notice it yet, as the most vulnerable people have already died by now. This happens, because the population is unable to select against virulence, because there is an antibody response against regions of the Spike protein that would normally not be targeted.
The previous failed vaccination experiment
I think this is just genuinely going to escalate dramatically.Why?
Well, we can look at the attempt to vaccinate chickens against influenza in Asia. These vaccines began selecting new variants, they ended up breeding versions of influenza that are extremely deadly and focus on infecting the brain rather than the lungs. Those versions have now spread around the world and killed millions of wild birds, entire species have had more than half their population die in a single season.
SARS2 is happily proceeding along the same path, with vaccine resistant versions emerging that are getting better at infecting the brain and increasingly competent at fusing cells together. In both cases we’re dealing with a virus that has an abnormal polybasic cleavage site, allowing it to infect endothelial cells.
If we’ve tried the exact same experiment of vaccinating against rapidly mutating respiratory viruses in another species and ended up breeding deadly viruses that decimate wild bird populations, wouldn’t you expect that performing the same experiment in our own species also results in deadly viruses that decimate our population?
You might argue to me that people now have immunity against proteins other than Spike too. But I don’t think it’s going to be very relevant, because you can only neutralize viral particles through Spike.
Immunity from interfering defective viral particles
So what about the unvaccinated? Well, I have explained most of the puzzle a few times.But there’s one element I’ve never addressed: interfering defective viral particles. This is not difficult to understand. When a cell is infected with the virus and does not get killed, but tries to get rid of the virus on its own, it will contain defective viral proteins or viral RNA. When it then gets infected again later on with a new variant, it will produce defective viral particles, that stimulate the immune system without being able to complete the viral cycle. These defective particles interfere in the normal particles replicating.
NK cells are aggressive upon a first infection, destroying infected cells. Upon subsequent infections however, they tend to prefer telling infected cells to solve the problem themselves. They have receptors they can use, to detect large chunks of weird RNA, which they will then delete. This means you end up with some persistent broken RNA in those cells, only activated again upon reinfection.
That’s a normal part of how immunity is supposed to work. It has a small protective effect on unrelated viruses, but a major protective effect on closely related viruses. Once you vaccinate people, you tend to break this immune mechanism, as the cells capable of producing defective viral particles will get killed.
Once you make the antibodies and T cells do the job of dealing with this virus, you set the immune system up for failure, relying entirely on these two mechanisms that are very vulnerable to the effects of mutations to the Spike protein. Such failure could come very suddenly and unpredictably, as changes to cysteine bridges can cause the whole NTD to fold differently, thereby evading most antibodies.
So all these studies, arguing that the virus never leaves your body, are not discovering something shocking. They’re just discovering how the immune system works: It encourages infected cells to store broken genetic material of viruses, that interferes with their replication next time the virus shows up.
Sometimes this means the virus fails to spread in your own body, sometimes it means you pass on versions of a virus with reduced fitness. Vaccinate everyone with inactivated vaccines and you break this mechanism.
Solving the problem
The tragedy is that the immune reprogramming after vaccination is effectively permanent, it is merely recalled with every subsequent infection. The antibodies that bind to new versions of the Spike protein prohibit NK cells and other elements of the innate immune system from learning to properly do their job during subsequent infections. You can look at the antibody response to the newest JN.1 strain here:Adults with two shots of mRNA:
Unvaccinated adults:
The immune system is still stuck with an abnormally elevated antibody response to this virus. As a result, the virus continues to be under pressure to shorten its immunogenic NTD loops and increase its fusogenicity.
In contrast, the innate immune system will have to do the job among unvaccinated adults. These are mechanisms that select against virulence.
The situation we’re dealing with, is one where the intrinsic virulence of this virus will continue to rise, until the antibody pressure exerted by the vaccinated is diminished.
Everything I’m saying here is a natural outgrowth of how the immune system works.
The main purpose of antibodies is to exert selection against virulence associated epitopes. That’s why survivors of the 1918 influenza pandemic have antibodies that react with the 1918 pandemic, but not with modern influenza strains. These people who built up natural immunity, with antibodies against select virulence-associated epitopes, are the reason the 1918 strain of influenza was replaced over time by more benign strains: Their immune systems would only pass on more benign versions of influenza.
We broke that mechanism for our own short-term self-interest and made a handful of people billionaires in the process.
Last edited:
Heliobas Disciple
TB Fanatic
SOME OF THE COMMENTS FROM THE ARTICLE, I COPIED OVER THE ONES RADAGAST (THE AUTHOR) ANSWERED. There were also some back and forth about lab leak or naturally occuring, Radagast thinks lab leak but not really relevant to the discussion so I didn't copy that over. Feel free to go to the link to read all the comments.
~~~~~~
LSWM Lives Matter
April 16, 2024 at 1:22 pm
Thanks for another update, I’ll leave a more detailed comment later on today when I have more time, but for now, with regards to the following quote:
“So all these studies, arguing that the virus never leaves your body, are not discovering something shocking. They’re just discovering how the immune system works: It encourages infected cells to store broken genetic material of viruses, that interferes with their replication next time the virus shows up.”
This is a relief. I remember Kareninca was referencing a doctor (sorry can’t remember his name) who was claiming that there were viral reservoirs in the bodies of not just the vaccinated (which would make sense considering that IgG4 antibodies encourage chronic/persistent infections) BUT ALSO IN THE UNVACCINATED.
So hopefully this “interfering defective viral particles” phenomenon/mechanism that you described debunks the claims of that doctor.
Radagast
April 16, 2024 at 2:38 pm
Oh I’m sure there are unvaccinated people with persistent infections. A persistent infection is just an infection that lasts for a long time. But it’s a rapidly mutating RNA virus. Within any single individual, it rapidly loses fitness. It rapidly tends to lose its furin cleavage site for example, without person-to-person transmission.
Eventually, unless you’re severely immunocompromised (in which case you have more pressing concerns), such persistent infections decay into defective interfering viral particles, that are only activated during a reinfection.
If we had not vaccinated and had not practiced social distancing, we would have seen this phenomenon take place on a global scale: A gradual decay in the intrinsic fitness of the virus, as chunks of RNA are irreversibly lost, the signal drowning in the noise.
After all, MERS and SARS1 don’t just keep spreading around the globe forever either.
Info
April 16, 2024 at 5:24 pm
How much will the world population decline?
Radagast
April 16, 2024 at 6:07 pm
That depends on the timeframe we’re talking about.
PBridger
April 16, 2024 at 7:34 pm
Thanks for simplifying not so easy to comprehend science.
I am curious on your take for the mRNA injections they are creating for cancers. Do you think the original mRNA injections taken by those with cancer will help or hurt the antigens they are producing for the various cancer types?
Radagast
April 16, 2024 at 11:59 pm
When you develop cancer, your own immune system has to fight the cancer. In the vast majority of cases, it succeeds at that quite well, so you never find out you had (pre)cancerous cells. Some cancers are kept under control so long that you actually die of something else (commonly seen with prostate cancer).
Sometimes you’ll need autoantibodies, antibodies targeting your own proteins, to tag these cancerous cells so your immune system knows they need to be killed.
The problem is that IgG4 antibodies don’t instruct the immune system to kill a cell when they bind to it. In fact it’s even worse than that, they interfere with other antibodies binding to those same cell, they stop those other antibodies from properly instructing the immune system to kill a cell.
So that’s basically the main reason why there’s a worry that the IgG4 antibody response seen after mRNA vaccination could result in rapidly progressing cancer.
I know there are more esoteric mechanisms proposed as well, but this to me seems the main concern.
How big of a problem this is would need to be studied. But studying it would first require acknowledging that something went wrong and we’re still stuck trying to get the system to acknowledge that simple fact.
Keith
April 16, 2024 at 9:42 pm
If your scenario is true (and it seems a lot more plausible to me than Geert’s hivicron doom) then the medical community is going to come up with a 4-in-1 mRNA supervax targeting the 4 circulating known serotypes, right?
Which I suppose will lead to further immune escape and more serotypes.
We never should have gotten on this treadmill. I had a very bad feeling about it from the beginning.
Radagast
April 17, 2024 at 12:03 am
These new strains of the virus have decreasing immunogenicity, a logical consequence of evolution. Trying to meaningfully raise antibody titers to four serotypes would presumably lead to a lot autoimmune problems.
~~~~~~
LSWM Lives Matter
April 16, 2024 at 1:22 pm
Thanks for another update, I’ll leave a more detailed comment later on today when I have more time, but for now, with regards to the following quote:
“So all these studies, arguing that the virus never leaves your body, are not discovering something shocking. They’re just discovering how the immune system works: It encourages infected cells to store broken genetic material of viruses, that interferes with their replication next time the virus shows up.”
This is a relief. I remember Kareninca was referencing a doctor (sorry can’t remember his name) who was claiming that there were viral reservoirs in the bodies of not just the vaccinated (which would make sense considering that IgG4 antibodies encourage chronic/persistent infections) BUT ALSO IN THE UNVACCINATED.
So hopefully this “interfering defective viral particles” phenomenon/mechanism that you described debunks the claims of that doctor.
Radagast
April 16, 2024 at 2:38 pm
Oh I’m sure there are unvaccinated people with persistent infections. A persistent infection is just an infection that lasts for a long time. But it’s a rapidly mutating RNA virus. Within any single individual, it rapidly loses fitness. It rapidly tends to lose its furin cleavage site for example, without person-to-person transmission.
Eventually, unless you’re severely immunocompromised (in which case you have more pressing concerns), such persistent infections decay into defective interfering viral particles, that are only activated during a reinfection.
If we had not vaccinated and had not practiced social distancing, we would have seen this phenomenon take place on a global scale: A gradual decay in the intrinsic fitness of the virus, as chunks of RNA are irreversibly lost, the signal drowning in the noise.
After all, MERS and SARS1 don’t just keep spreading around the globe forever either.
Info
April 16, 2024 at 5:24 pm
How much will the world population decline?
Radagast
April 16, 2024 at 6:07 pm
That depends on the timeframe we’re talking about.
PBridger
April 16, 2024 at 7:34 pm
Thanks for simplifying not so easy to comprehend science.
I am curious on your take for the mRNA injections they are creating for cancers. Do you think the original mRNA injections taken by those with cancer will help or hurt the antigens they are producing for the various cancer types?
Radagast
April 16, 2024 at 11:59 pm
When you develop cancer, your own immune system has to fight the cancer. In the vast majority of cases, it succeeds at that quite well, so you never find out you had (pre)cancerous cells. Some cancers are kept under control so long that you actually die of something else (commonly seen with prostate cancer).
Sometimes you’ll need autoantibodies, antibodies targeting your own proteins, to tag these cancerous cells so your immune system knows they need to be killed.
The problem is that IgG4 antibodies don’t instruct the immune system to kill a cell when they bind to it. In fact it’s even worse than that, they interfere with other antibodies binding to those same cell, they stop those other antibodies from properly instructing the immune system to kill a cell.
So that’s basically the main reason why there’s a worry that the IgG4 antibody response seen after mRNA vaccination could result in rapidly progressing cancer.
I know there are more esoteric mechanisms proposed as well, but this to me seems the main concern.
How big of a problem this is would need to be studied. But studying it would first require acknowledging that something went wrong and we’re still stuck trying to get the system to acknowledge that simple fact.
Keith
April 16, 2024 at 9:42 pm
If your scenario is true (and it seems a lot more plausible to me than Geert’s hivicron doom) then the medical community is going to come up with a 4-in-1 mRNA supervax targeting the 4 circulating known serotypes, right?
Which I suppose will lead to further immune escape and more serotypes.
We never should have gotten on this treadmill. I had a very bad feeling about it from the beginning.
Radagast
April 17, 2024 at 12:03 am
These new strains of the virus have decreasing immunogenicity, a logical consequence of evolution. Trying to meaningfully raise antibody titers to four serotypes would presumably lead to a lot autoimmune problems.
Zoner
Veteran Member
Thanks HD. He has reached the same conclusions as Geert and Nikhil. And as you say a different scientific explanation and yet the same in structure is offered. Two witnesses confirm a thing and ....... so it begins.IF YOU ONLY HAVE TIME TO READ ONE POST I POSTED TODAY, READ THIS ONE.
You don't have to understand all the science (I surely don't), but keep reading past what you don't understand so you get the general picture. Radagast is reaching the same conclusion Geert reaches, although I'm not sure if their way to get to that conclusion is the same because the science is over my head and I don't have the patience to try to figure it out like I used to do with Geert (I don't try anymore with him either, I realized after hours listening and figuring out the science after a lot of concentration that it doesn't matter if I follow it all to get the gist of what they're saying). It's worth reading even if we don't understand all of the nitty gritty. I would love if Phillip McMillan would interview both of them together and get a discussion going! (and break it down so we can all understand all of it)
~~~~~~~~~~~~~~~~~
(fair use applies)
Hey look, serotypes!
Radagast
April 16, 2024
I have to point this out. About half a year ago, I said that you’re probably going to end up with SARS2 serotypes, simply because it was unlikely by now that something new that emerges would be superior enough to completely eradicate the XBB lineages. Immunity would first be reoriented against the new serotype and that would then create a landscape within which the old serotype could re-emerge.
Well guess what happened. It turns out there’s an XBB lineage with a weird insertion at 212 that emerged in Canada and spread around the globe. So now you have two versions of the virus, both with insertions in different parts of the NTD. They’re both going to be under pressure to change the NTD in different ways. For JN.1 it’s straightforward to start deleting stuff around the first loop, due to the insertion at 16. We see it happening in multiple JN.1 lineages, a deletion at 31, just after the first immunogenic loop and a bit after the insertion, that allows a new glycan to be added at 30. But this XBB variant would have more of an incentive to delete chunks further along in the NTD.
Remember, it’s when deletions and new glycans emerge in the NTD, that the last antibodies that neutralize the virus in most of the population are going to fail. You will end up with very fusogenic variants, that spread by moving from one cell into another, without ever being in reach of the antibodies. The antibody response against the RBD is already pretty much useless by now.
The important thing to keep in mind is that once you have multiple of these highly fusogenic serotypes circulating in the population, you can’t vaccinate anymore. Most of the population already has high concentrations of antibodies, vaccination under these conditions won’t improve the population’s protection, it will merely favor one serotype at the cost of the other. By now it’s clear that the BA.2.86* Spike doesn’t do a clean sweep: It stays stuck around 93%, so it doesn’t completely wipe out the other lineages.
There are probably other serotypes out there too, we just don’t see them because sequencing is down by 99%. There’s a weird one in France, but nobody knows how widespread it is. There’s also a derivative of the original Omicron still circulating in Nebraska, infecting multiple people.
I know you’ve probably seen it argued there will be some sudden new doomsday variant this summer that leads to overnight vaccine failure. I don’t think this is likely, although it is inherently impossible to rule out.
In the coming weeks we can expect S:R346T, S:F456L, S:S31del to grow globally dominant.
Right now, adding the whole thing together suggests a 117% growth advantage.
What seems far more likely to me unfortunately, is that we’re gradually going to see different serotypes emerge: XBB lineages and BA.2.86 lineages, along with lineages based on whatever else is still out there.
Those lineages will undergo deletions in the immunogenic loops of the N-Terminal Domain, facilitated by the rare insertions they tend to develop in the NTD. These deletions will increase fusogenicity and thereby increase the intrinsic virulence of the virus.
That ultimately results in a virus that behaves more like the original SARS.
The reason this is happening
So why did this happen? Well, allow me to show you something.
There is a notable difference by now between vaccinated and unvaccinated people, in regards to the regions their T cell receptors look for. This has been studied, in people who were infected in the first half of 2022.
Among the unvaccinated, they found strong overrepresentation of the S673-699 epitope. On the other hand, among the vaccinated, they found strong overrepresentation of these four regions:
S135-177, S264-276, S319-350, and S448-472.
The first of these four is easy to understand: It’s the N3 loop of the NTD. The other three regions are also in the NTD, but don’t correspond neatly to any of the known immunogenic loops.
But here’s the whole picture, which tells the story in my opinion:
I would argue it’s pretty obvious what’s going on here. The unvaccinated adaptive immune response tends to focus on regions in S2. The vaccinated adaptive immune response tends to focus on the N-Terminal Domain. We also see it strongly preferentially focuses on the region around 180, which is an infection enhancing site. The N5 loop also seems to be preferred.
So it seems straightforward, to expect that the NTD is going to undergo radical changes now, now that antibodies against the RBD have become useless. As the loops in the NTD grow shorter, by deletion mutations, you end up with a virus that behaves more like SARS1.
The entire Spike protein is covered by glycans, with the exception of the receptor binding domain. But now that the RBD evades the antibodies, we’re reaching the point where the antibody response depends on those last few regions of the NTD not covered by glycans, where antibodies can still bind to neutralize viral particles.
You also have to keep in mind: As more and more of the Spike protein becomes inaccessible to the T-cells, whether due to evolution to resemble our own proteins, deletions, steric hindrance, or glycans that are added, you’re going to see people’s lungs populated by an increasingly homogeneous population of T-cells.
With every breakthrough infection, the immunological abnormalities further escalate. That’s why you now see a very different antibody response against JN.1, when comparing vaccinated and unvaccinated people, as the vaccinated keep recalling antibodies against recessive epitopes in the NTD. This is what leads to the current situation, in which people suffer constantly elevated levels of respiratory disease. A lung environment populated with T cells looking for SARS2 has little room for other T cells looking for unrelated respiratory pathogens.
Through vaccination, you put abnormal antibody pressure on the Spike protein. This abnormal antibody pressure leads to affinity escape (escaping low affinity antibodies by increasing ACE2 affinity). But now that the abnormal antibody pressure is so strongly concentrated on the NTD, you’re encouraging the immunogenic loops of the NTD to grow shorter, which is known to dramatically increase fusogenicity. In other words, you are selecting more virulent variants.
It’s very simple: You want to see a small number of high affinity antibodies, against select regions of the Spike protein, in people who suffered severe infections. Those antibodies will be against regions that are associated with virulence. As a result, people will gradually offer a selective advantage to variants of this virus that are less virulent. What you don’t want to see, is this yellow line in the whole population:
This is what happened to the four hCovs in our species: We had a pandemic long ago (including one in the 19th century), a bunch of people died, then it gradually grew less virulent. But here’s what’s happening to SARS2:
It has been gradually growing more virulent since the first Omicron, but we don’t really notice it yet, as the most vulnerable people have already died by now. This happens, because the population is unable to select against virulence, because there is an antibody response against regions of the Spike protein that would normally not be targeted.
The previous failed vaccination experiment
I think this is just genuinely going to escalate dramatically.
Why?
Well, we can look at the attempt to vaccinate chickens against influenza in Asia. These vaccines began selecting new variants, they ended up breeding versions of influenza that are extremely deadly and focus on infecting the brain rather than the lungs. Those versions have now spread around the world and killed millions of wild birds, entire species have had more than half their population die in a single season.
SARS2 is happily proceeding along the same path, with vaccine resistant versions emerging that are getting better at infecting the brain and increasingly competent at fusing cells together. In both cases we’re dealing with a virus that has an abnormal polybasic cleavage site, allowing it to infect endothelial cells.
If we’ve tried the exact same experiment of vaccinating against rapidly mutating respiratory viruses in another species and ended up breeding deadly viruses that decimate wild bird populations, wouldn’t you expect that performing the same experiment in our own species also results in deadly viruses that decimate our population?
You might argue to me that people now have immunity against proteins other than Spike too. But I don’t think it’s going to be very relevant, because you can only neutralize viral particles through Spike.
Immunity from interfering defective viral particles
So what about the unvaccinated? Well, I have explained most of the puzzle a few times.
But there’s one element I’ve never addressed: interfering defective viral particles. This is not difficult to understand. When a cell is infected with the virus and does not get killed, but tries to get rid of the virus on its own, it will contain defective viral proteins or viral RNA. When it then gets infected again later on with a new variant, it will produce defective viral particles, that stimulate the immune system without being able to complete the viral cycle. These defective particles interfere in the normal particles replicating.
NK cells are aggressive upon a first infection, destroying infected cells. Upon subsequent infections however, they tend to prefer telling infected cells to solve the problem themselves. They have receptors they can use, to detect large chunks of weird RNA, which they will then delete. This means you end up with some persistent broken RNA in those cells, only activated again upon reinfection.
That’s a normal part of how immunity is supposed to work. It has a small protective effect on unrelated viruses, but a major protective effect on closely related viruses. Once you vaccinate people, you tend to break this immune mechanism, as the cells capable of producing defective viral particles will get killed.
Once you make the antibodies and T cells do the job of dealing with this virus, you set the immune system up for failure, relying entirely on these two mechanisms that are very vulnerable to the effects of mutations to the Spike protein. Such failure could come very suddenly and unpredictably, as changes to cysteine bridges can cause the whole NTD to fold differently, thereby evading most antibodies.
So all these studies, arguing that the virus never leaves your body, are not discovering something shocking. They’re just discovering how the immune system works: It encourages infected cells to store broken genetic material of viruses, that interferes with their replication next time the virus shows up.
Sometimes this means the virus fails to spread in your own body, sometimes it means you pass on versions of a virus with reduced fitness. Vaccinate everyone with inactivated vaccines and you break this mechanism.
Solving the problem
The tragedy is that the immune reprogramming after vaccination is effectively permanent, it is merely recalled with every subsequent infection. The antibodies that bind to new versions of the Spike protein prohibit NK cells and other elements of the innate immune system from learning to properly do their job during subsequent infections. You can look at the antibody response to the newest JN.1 strain here:
Adults with two shots of mRNA:
Unvaccinated adults:
The immune system is still stuck with an abnormally elevated antibody response to this virus. As a result, the virus continues to be under pressure to shorten its immunogenic NTD loops and increase its fusogenicity.
In contrast, the innate immune system will have to do the job among unvaccinated adults. These are mechanisms that select against virulence.
The situation we’re dealing with, is one where the intrinsic virulence of this virus will continue to rise, until the antibody pressure exerted by the vaccinated is diminished.
Everything I’m saying here is a natural outgrowth of how the immune system works.
The main purpose of antibodies is to exert selection against virulence associated epitopes. That’s why survivors of the 1918 influenza pandemic have antibodies that react with the 1918 pandemic, but not with modern influenza strains. These people who built up natural immunity, with antibodies against select virulence-associated epitopes, are the reason the 1918 strain of influenza was replaced over time by more benign strains: Their immune systems would only pass on more benign versions of influenza.
We broke that mechanism for our own short-term self-interest and made a handful of people billionaires in the process.
Tristan
Has No Life - Lives on TB
Dr. Campbell and Dr. Dalgliesh discuss their concerns regarding the increases in cancers
View: https://www.youtube.com/watch?v=6H0bChkxCWs
rt: 15:33
rt: 15:33
Heliobas Disciple
TB Fanatic
Biden administration announces new partnership with 50 countries to stifle future pandemics
U.S. government officials will offer support in the countries to develop better preparedness for such outbreaks in those countries.
 apnews.com
apnews.com
Biden administration announces new partnership with 50 countries to stifle future pandemics
By AMANDA SEITZ
April 17, 2024
WASHINGTON (AP) — President Joe Biden’s administration will help 50 countries identify and respond to infectious diseases, with the goal of preventing pandemics like the COVID-19 outbreak that suddenly halted normal life around the globe in 2020.
U.S. government officials will offer support in the countries, most of them located in Africa and Asia, to develop better testing, surveillance, communication and preparedness for such outbreaks in those countries.
The strategy will help “prevent, detect and effectively respond to biological threats wherever they emerge,” Biden said in a statement Tuesday.
The Global Health Security Strategy, the president said, aims to protect people worldwide and “will make the United States stronger, safer, and healthier than ever before at this critical moment.”
The announcement about the strategy comes as countries have struggled to meet a worldwide accord on responses to future pandemics. Four years after the coronavirus pandemic, prospects of a pandemic treaty signed by all 194 of the World Health Organization’s members are flailing.
Talks for the treaty are ongoing, with a final text expected to be agreed upon next month in Geneva. It’s meant to be a legally binding treaty that obliges countries to monitor pandemic threats and share scientific findings. But major disputes have emerged over vaccine equity and transferring the technology used to make vaccines.
Even if a deal is hammered out, there would be few consequences for countries that choose not to abide by the treaty.
The U.S. will push on with its global health strategy to prevent future pandemics, regardless of a pandemic treaty or not, a senior administration official told reporters on Monday.
Several U.S. government agencies — including the State Department, the Centers for Disease Control and Prevention, Health and Human Services and the U.S. Agency for International Development, or USAID — will help countries refine their infectious disease response.
Health systems around the globe have been overwhelmed with COVID-19 and other health emergencies such as Ebola, malaria and mpox, the CDC said in a statement. The new strategy will help countries rebuild their agencies, the U.S. agency explained.
“Global health security is national security, and CDC is proud to contribute its expertise, investments and rapid response to protect the health and safety of the American people and the world,” Mandy Cohen, the CDC’s director, said in a statement.
Congo is one country where work has already begun. The U.S. government is helping Congo with its response to an mpox virus outbreak, including with immunizations. Mpox, a virus that’s in the same family as the one that causes smallpox, creates painful skin lesions. The World Health Organization declared mpox a global emergency in 2022, and there have been more than 91,000 cases spanning across 100 countries to date.
The White House on Tuesday released a website with the names of the countries that are participating in the program. Biden officials are seeking to get 100 countries signed onto the program by the end of the year.
The U.S. has devoted billions of dollars, including money raised from private donations, to the effort. Biden, a Democrat, is asking for $1.2 billion for global health safety efforts in his yearly budget proposal to Congress.
Heliobas Disciple
TB Fanatic

USDA funding $1M study with China to make bird flu more contagious
The US government is spending $1million of American taxpayer money to fund dangerous experiments with bird flu viruses in collaboration with Chinese scientists.
Chinese scientists to make bird flu strains more infectious and deadly as part of $1m project - despite fears similar tests unleashed Covid
By Alexa Lardieri U.S. Deputy Health Editor Dailymail.Com
Published: 18:35 EDT, 15 February 2024 | Updated: 14:43 EDT, 16 February 2024
- Research involves infecting birds with flu viruses to make them more infectious
- The USDA is spending more than $1million to fund the risky experiments
The US government is spending $1million of American taxpayer money to fund experiments on dangerous bird flu viruses in collaboration with Chinese scientists.
The research involves infecting ducks and geese with different strains of viruses to make them more infectious, and study the viruses' potential to 'jump into mammalian hosts,' according to research documents.
It is being funded through the US Department of Agriculture and collaborating studies will take place at sites in Georgia, Beijing and Edinburgh in Scotland.
It comes despite similar research being restricted in 2022 and growing concerns that dubious Chinese studies may have started the Covid pandemic.
The documents were obtained by the campaign group, The White Coat Waste Project, and shared with DailyMail.com.
The papers show funding for the avian virus research began in April 2021 and it is slated to continue through March 2026.
The USDA told this website the project was applied for in 2019 and approved in 2020.
The specific viruses the researchers will work with include H5NX, H7N9 and H9N2, WCW reported.
A 2023 study described H5NX viruses as 'highly pathogenic' with the ability to cause neurological complications in humans.
The H7N9 strain first infected humans and animals in China in March 2013 and the World Health Organization said it is of concern 'because most patients have become severely ill.'
The H9N2 strain has been found in dove in China and while it has a lower pathogenicity than the other strains, it can still infect humans.
The main collaborators on the project are USDA Southeast Poultry Research Laboratory, the Chinese Academy of Sciences, and the University of Edinburgh’s Roslin Institute - a Wuhan lab partner.
Additionally, one of the researchers collaborating on the project is Wenju Liu, who is affiliated with the WIV - which is believed to have sparked the Covid pandemic - and a member of the board of a scientific journal, working with Zheng-Li Shi, who is known as the 'bat lady' for her extensive work on bat coronaviruses.
Different aspects of the research are slated to take place in multiple locations, including poultry research centers in Athens, Georgia, at the Roslin Institute in Edinburgh and at the Chinese academy in Beijing.
Allan Rodriguez, a spokesperson with the USDA, told DailyMail.com the $100million funding awarded to the research 'is only being committed to the specific components carried out by our own team located in Athens, Georgia, and is not any way contributing to research taking place in the UK or China.'
Justin Goodman, senior vice president of the WCW, said in a statement to DailyMail.com: 'It's reckless and indefensible for... bureaucrats to bankroll dangerous avian flu gain-of-function studies involving virus experimenters from the notorious Wuhan animal lab that likely caused COVID and its CCP-run parent organization, the Chinese Academy of Sciences.
'Taxpayers shouldn’t be forced to foot the bill for animal experiments with foreign adversaries that soup up viruses and can cause pandemics or create bioweapons.'
Following the publication of the documents by WCW, Republican Sen Joni Ernst of Iowa wrote a letter Thursday to Tom Vilsack, secretary of the USDA, seeking more information about the department's ongoing funding of the research.
The letter read: 'I was troubled to learn from the non-profit group White Coat Waste Project that USDA is supporting experiments involving a "highly pathogenic avian influenza virus" that poses a "risk to both animals and humans."'
Sen Ernst said in a statement to DailyMail.com: 'The health and safety of Americans are too important to just wing it, and Biden’s USDA should have had more apprehension before sending any taxpayer dollars to collaborate with [China] on risky avian flu research.
'They should know by now to suspect "fowl" play when it comes to researchers who have ties to the dangerous Wuhan Lab, and simply switching from bats to birds causes concern that they are creating more pathogens of pandemic potential.
'Here’s my warning: the Biden administration should be walking on eggshells until they cut off every cent going to our adversaries. We cannot allow what happened in Wuhan to happen again.'
Despite the senators concerns, the USDA spokesperson told this website it is 'common for international researchers to conduct independent research that's connected to the same end goal' and that the research does not qualify as gain-of-function.
He added: 'Any time Senator Ernst has a question for USDA about our research or our commitment to helping America’s farmers mitigate animal diseases like highly pathogenic avian influenza, she should reach out to us directly before putting misinformation in a press release or public letter... and despite the senator's assertions, this is not gain-of-function research.
'Because animal diseases present a global threat, it is common for international researchers to conduct independent research that’s connected to the same end goal – but what Senator Ernst lays out in her letter is far off base from what’s actually transpiring, and on top of that is based on approval decisions that predate this Administration.'
Heliobas Disciple
TB Fanatic

Fury as it's revealed US and Chinese lab are creating deadly viruses
Lawmakers are demanding answers after it was revealed the USDA is supplying China with money for experiments that see scientists make bird flu viruses more contagious to humans.
USDA and China CCP lab are creating deadly BIRD FLU viruses as part of $1m collaboration - and YOU are paying for it
By Alexa Lardieri U.S. Deputy Health Editor Dailymail.Com
Published: 07:32 EDT, 17 April 2024 | Updated: 09:15 EDT, 17 April 2024
- A letter states the research could threaten national security and public health
- It involves infecting ducks with different viruses to make them more infectious
- DailyMail.com asked experts if H5N1 case in Texas raises risk of future pandemic
Lawmakers are demanding answers after it was revealed the US is sending taxpayer dollars to a Chinese army lab to make bird flu viruses more dangerous to people.
Eighteen members of Congress are demanding answers from the Department of Agriculture (USDA) about the project, which was first revealed by DailyMail.com.
It is part of a $1million collaboration between the USDA and the CCP-run Chinese Academy of Sciences - the institution that oversees the Wuhan lab at the center of the Covid lab-leak theory.
In a scathing letter to USDA Secretary Tom Vilsack last week, the bipartisan group said: 'This research, funded by American taxpayers, could potentially generate dangerous new lab-created virus strains that threaten our national security and public health.'
The research comes as fears about bird flu rise. A farm worker in Texas caught the H5N1 strain that is racing through cattle across the US earlier this month, becoming only the second ever American to be diagnosed - and experts are bracing for more cases.
In February, it was revealed the United States government was funneling $1million to China to see if scientists could make 'highly pathogenic avian influenza' more contagious to mammals using gain-of-function research.
Government records showed the collaboration began in April 2021 and is scheduled to be funded through March 2026. The USDA previously told this website the project was applied for in 2019 and approved in 2020.
The research involves infecting ducks and geese with different strains of viruses to make them more infectious, and studying the viruses' potential to 'jump into mammalian hosts,' according to research documents obtained by the watchdog group White Coat Waste Project and shared exclusively with this website.
It is being funded through the USDA and the main collaborators on the project are USDA Southeast Poultry Research Laboratory, the Chinese Academy of Sciences and the University of Edinburgh’s Roslin Institute - a Wuhan lab partner.
And it has been ongoing despite similar research being restricted in 2022 and growing concerns that dubious Chinese studies may have started the Covid pandemic.
Just yesterday, President Joe Biden's administration announced it will work with 50 countries to identify and respond to infectious diseases, with the goal of preventing a pandemic that the US' own research could actually spark.
Last week's letter was spearheaded by Rep Nick Langworthy, a Republican from New York who serves on the House Agriculture Committee.
It states: 'We are disturbed by recent reports about the US Department of Agriculture’s (USDA) collaboration with the Chinese Communist Party (CCP)-linked Chinese Academy of Sciences (CAS) on bird flu research.'
The CAS is the parent organization of WIV and has previously been prohibited from receiving US government money for 'blatantly violating grant and biosafety policies, refusing to share lab notebooks and other data, and otherwise obstructing investigations into the likely role of the lab's risky coronavirus [gain of function] research in the origin of Covid-19.'
The letter continued: 'Recognizing the problematic behavior of CAS, our House and Senate colleagues have called for sanctions against CAS and its affiliates and for taxpayer funding to be cut for all research involving CAS.'
The signatories then requested written answers to seven questions inquiring about the potential of the research to increase transmissibility of bird flu viruses, details about specific experiments being performed, the biosafety levels of the experiments, what oversight the USDA is providing over CAS and if the FBI performed a safety risk assessment on the collaboration - and, if so, what those results were.
Bird flu is of particular concern right now because a farmer in Texas recently contracted the H5N1 strain of the virus.
The patient caught the bird flu from an infected cow, which was the first time the strain had been found in cattle.
They are only the second person to contract H5N1 after someone in Colorado caught the virus in 2022.
While there is no sign of person-to-person spread — a development that would signal the start of a human epidemic — experts say the ease with which the strain is jumping between species raises the risk of it evolving to infect people more easily.
The Centers for Disease Control and Prevention report a 'low' public health risk.
The virus, however, is widespread in wild birds, with sporadic infections in poultry and mammals.
Experts have previously told DailyMail.com H5N1 has the potential to spark a new pandemic.
Dr Aaron Glatt, an infectious diseases expert at Mount Sinai in New York, warned: 'It is absolutely true that H5N1 has the potential to cause a pandemic.
'People who work with these animals do need to be careful.
'The more that this virus is spread, the more likely it is that it could become a strain that could mutate and start to spread from human-to-human.'
The H5N1 spreading across the world emerged in 2020 after a bird was infected with both a bird flu from domestic poultry and a virus from wild birds.
During the infection, the two viruses met in the same cell and swapped genes — in a process scientifically termed re-assortment — to create the new virus that now had multiple attributes that made it better at infecting bird cells.
It quickly spread globally, with the first cases identified in Europe — before infections were also detected in Africa, the Middle East and Asia.
This month's letter is not the first written to the USDA from lawmakers.
Following February's investigation, Republican Sen Joni Ernst of Iowa wrote a letter to Sec Vilsack seeking more information about the department's ongoing funding of the research.
The letter read: 'I was troubled to learn from the non-profit group White Coat Waste Project that USDA is supporting experiments involving a "highly pathogenic avian influenza virus" that poses a "risk to both animals and humans."'
Sen Ernst said in a statement to DailyMail.com at the time: 'The health and safety of Americans are too important to just wing it, and Biden’s USDA should have had more apprehension before sending any taxpayer dollars to collaborate with [China] on risky avian flu research.
'They should know by now to suspect "fowl" play when it comes to researchers who have ties to the dangerous Wuhan Lab, and simply switching from bats to birds causes concern that they are creating more pathogens of pandemic potential.
'Here’s my warning: the Biden administration should be walking on eggshells until they cut off every cent going to our adversaries. We cannot allow what happened in Wuhan to happen again.'
The specific viruses the research said it will study include H5NX, H7N9 and H9N2, WCW reported.
A 2023 study described H5NX viruses as 'highly pathogenic' with the ability to cause neurological complications in humans.
The H7N9 strain first infected humans and animals in China in March 2013 and the World Health Organization said it is of concern 'because most patients have become severely ill.'
The H9N2 strain has been found in doves in China and while it has a lower pathogenicity than the other strains, it can still infect humans.
Despite the concerns, a USDA spokesperson told this website it is 'common for international researchers to conduct independent research that's connected to the same end goal' and that the research does not qualify as gain-of-function.
Heliobas Disciple
TB Fanatic
(fair use applies)
H5N1 Circulating In Dairy Cows And Cats In America Are A New Strain: B3.13 Belonging To Clade 2.3.4.4b That Underwent Reassortment Events
Nikhil Prasad Fact checked by:Thailand Medical News Team
Apr 18, 2024
Highly pathogenic avian influenza (HPAI) A (H5N1) viruses belonging to clade 2.3.4.4b have emerged as a significant global public health threat, impacting poultry, wildlife, and potentially humans. Recent developments in the United States have revealed a new strain, genotype B3.13, within this clade, affecting domestic dairy cattle and domestic cats. This H5N1 News report delves into the origins, detection, genomic characterization, phylogenetic analysis, mutation adaptations, and potential implications of this novel HPAI virus strain.
Emergence and Spread of Clade 2.3.4.4b H5N1 Virus
The clade 2.3.4.4b H5N1 virus originated in East and Southeast Asia in 2020, primarily affecting avian species. Its subsequent spread globally via migratory birds and trade networks led to its introduction to North America in late 2021, followed by infections in wild terrestrial mammals across the continent. The Texas Panhandle, situated within major bird migratory flyways, experienced an outbreak in dairy cattle and domestic cats in early 2024, signaling a new phase of the virus's spread and adaptation.
Detection and Genomic Characterization
In February and March 2024, dairy cattle in Texas exhibited symptoms such as reduced feed intake and decreased milk production, while domestic cats showed similar signs after consuming raw colostrum and milk. Samples from affected animals tested positive for HPAI H5N1 clade 2.3.4.4b, leading to genomic sequencing that identified genotype B3.13, indicating reassortment events involving HPAI and low pathogenic avian influenza (LPAI) viruses.
The genomic analysis revealed that the B3.13 genotype had undergone two reassortment events since 2023, leading to critical mutations in genes such as HA, M1, and NS. Notably, these mutations did not include key changes in PB2 and PB1 genes associated with enhanced virulence or adaptation to mammalian hosts. However, a human case with the PB2 E627K mutation highlighted the virus's potential for rapid evolution post-infection, necessitating continued surveillance.
Phylogenetic and Reassortment Analysis
Phylogenetic analysis revealed a close relationship among viruses from dairy cattle, domestic cats, wild birds, and a human case, forming a distinct subcluster within clade 2.3.4.4b. Reassortment events between HPAI and LPAI viruses contributed to genotype B3.13, highlighting the virus's evolving nature and adaptability across species barriers.
Further analysis using the GenoFlu tool classified the B3.13 genotype as a result of reassortment involving genotype B3.7 and an LPAI virus strain. The HA, NA, PA, and MP genes originated from the H5N1 virus strain A1 in 2020, while other segments were closely related to LPAI viruses. This intricate reassortment pattern underscores the virus's dynamic genetic makeup and its potential for rapid ev olution.
Critical Amino Acid Mutation Analysis
Amino acid mutation analysis identified key residues in the HA, M1, and NS1 genes associated with increased virulence and potential human receptor binding affinity. Notably, mutations linked to mammalian host adaptation and enhanced transmission were absent in HPAI viruses from dairy cattle and cats, mitigating immediate concerns for widespread human transmission.
Specifically, mutations in the HA gene residues 137A, 158N, and 160A were observed, which may increase binding affinity to human-type receptors. Additionally, mutations in M1 (30D, 43M, and 215A) and NS1 (42S, 103F, and 106M) were consistent with patterns observed in HPAI isolates from terrestrial and marine mammals, indicating potential virulence in mammalian hosts.
However, critical mutations associated with mammalian host adaptation and enhanced transmission, such as residues 591K, 627K/V/A, 701N in PB2, and 228S, were absent in all HPAI virus strains derived from dairy cattle and cats. This observation suggests that while the current risk to human health is relatively low, ongoing surveillance and vigilance are crucial due to the virus's potential for adaptive changes.
Implications and Public Health Concerns
The detection of HPAI H5N1 clade 2.3.4.4b in domestic dairy cattle raises concerns about zoonotic transmission through unpasteurized milk, necessitating public awareness and food safety measures. While current mutations pose a low immediate risk to human health, the virus's potential for rapid evolution underscores the need for continued surveillance and preparedness.
Additionally, the detection of the same strain of HPAI viruses in various wild bird species, such as blackbirds and common grackles in Texas and Canada geese in Wyoming (Central Flyway), provides further support for potential transmission routes. This includes direct contact or contamination of water sources and feed, highlighting the complex interplay between migratory birds, wildlife, and domestic animals in virus transmission dynamics.
Furthermore, the recent confirmation of this HPAI virus strain in dairy herds across multiple states emphasizes the need for coordinated response efforts and biosecurity measures within the agricultural sector. The interconnectedness of wildlife, livestock, and human populations necessitates a One Health approach to disease management, encompassing veterinary, public health, and environmental perspectives.
Conclusion
The emergence of HPAI H5N1 clade 2.3.4.4b in domestic dairy cattle and cats marks a significant development in the ongoing global influenza landscape. Understanding the virus's genomic characteristics, reassortment events, and mutation patterns is crucial for effective surveillance, outbreak management, and public health interventions to mitigate potential threats to human and animal populations. Continued research and collaboration across disciplines are essential in addressing the dynamic challenges posed by emerging infectious diseases like HPAI.
The study findings by researchers from Iowa State University-USA were published on a preprint server and are currently being peer reviewed.

 www.biorxiv.org
www.biorxiv.org
For the latest H5N1 News, keep on logging to Thailand Medical News.
Read Also:
https://www.thailandmedical.news/ne...ave-already-been-infected-with-the-new-strain
https://www.thailandmedical.news/ne...uza-do-not-work-against-the-new-h5n1-variants
H5N1 Circulating In Dairy Cows And Cats In America Are A New Strain: B3.13 Belonging To Clade 2.3.4.4b That Underwent Reassortment Events
Nikhil Prasad Fact checked by:Thailand Medical News Team
Apr 18, 2024
Highly pathogenic avian influenza (HPAI) A (H5N1) viruses belonging to clade 2.3.4.4b have emerged as a significant global public health threat, impacting poultry, wildlife, and potentially humans. Recent developments in the United States have revealed a new strain, genotype B3.13, within this clade, affecting domestic dairy cattle and domestic cats. This H5N1 News report delves into the origins, detection, genomic characterization, phylogenetic analysis, mutation adaptations, and potential implications of this novel HPAI virus strain.
Emergence and Spread of Clade 2.3.4.4b H5N1 Virus
The clade 2.3.4.4b H5N1 virus originated in East and Southeast Asia in 2020, primarily affecting avian species. Its subsequent spread globally via migratory birds and trade networks led to its introduction to North America in late 2021, followed by infections in wild terrestrial mammals across the continent. The Texas Panhandle, situated within major bird migratory flyways, experienced an outbreak in dairy cattle and domestic cats in early 2024, signaling a new phase of the virus's spread and adaptation.
Detection and Genomic Characterization
In February and March 2024, dairy cattle in Texas exhibited symptoms such as reduced feed intake and decreased milk production, while domestic cats showed similar signs after consuming raw colostrum and milk. Samples from affected animals tested positive for HPAI H5N1 clade 2.3.4.4b, leading to genomic sequencing that identified genotype B3.13, indicating reassortment events involving HPAI and low pathogenic avian influenza (LPAI) viruses.
The genomic analysis revealed that the B3.13 genotype had undergone two reassortment events since 2023, leading to critical mutations in genes such as HA, M1, and NS. Notably, these mutations did not include key changes in PB2 and PB1 genes associated with enhanced virulence or adaptation to mammalian hosts. However, a human case with the PB2 E627K mutation highlighted the virus's potential for rapid evolution post-infection, necessitating continued surveillance.
Phylogenetic and Reassortment Analysis
Phylogenetic analysis revealed a close relationship among viruses from dairy cattle, domestic cats, wild birds, and a human case, forming a distinct subcluster within clade 2.3.4.4b. Reassortment events between HPAI and LPAI viruses contributed to genotype B3.13, highlighting the virus's evolving nature and adaptability across species barriers.
Further analysis using the GenoFlu tool classified the B3.13 genotype as a result of reassortment involving genotype B3.7 and an LPAI virus strain. The HA, NA, PA, and MP genes originated from the H5N1 virus strain A1 in 2020, while other segments were closely related to LPAI viruses. This intricate reassortment pattern underscores the virus's dynamic genetic makeup and its potential for rapid ev olution.
Critical Amino Acid Mutation Analysis
Amino acid mutation analysis identified key residues in the HA, M1, and NS1 genes associated with increased virulence and potential human receptor binding affinity. Notably, mutations linked to mammalian host adaptation and enhanced transmission were absent in HPAI viruses from dairy cattle and cats, mitigating immediate concerns for widespread human transmission.
Specifically, mutations in the HA gene residues 137A, 158N, and 160A were observed, which may increase binding affinity to human-type receptors. Additionally, mutations in M1 (30D, 43M, and 215A) and NS1 (42S, 103F, and 106M) were consistent with patterns observed in HPAI isolates from terrestrial and marine mammals, indicating potential virulence in mammalian hosts.
However, critical mutations associated with mammalian host adaptation and enhanced transmission, such as residues 591K, 627K/V/A, 701N in PB2, and 228S, were absent in all HPAI virus strains derived from dairy cattle and cats. This observation suggests that while the current risk to human health is relatively low, ongoing surveillance and vigilance are crucial due to the virus's potential for adaptive changes.
Implications and Public Health Concerns
The detection of HPAI H5N1 clade 2.3.4.4b in domestic dairy cattle raises concerns about zoonotic transmission through unpasteurized milk, necessitating public awareness and food safety measures. While current mutations pose a low immediate risk to human health, the virus's potential for rapid evolution underscores the need for continued surveillance and preparedness.
Additionally, the detection of the same strain of HPAI viruses in various wild bird species, such as blackbirds and common grackles in Texas and Canada geese in Wyoming (Central Flyway), provides further support for potential transmission routes. This includes direct contact or contamination of water sources and feed, highlighting the complex interplay between migratory birds, wildlife, and domestic animals in virus transmission dynamics.
Furthermore, the recent confirmation of this HPAI virus strain in dairy herds across multiple states emphasizes the need for coordinated response efforts and biosecurity measures within the agricultural sector. The interconnectedness of wildlife, livestock, and human populations necessitates a One Health approach to disease management, encompassing veterinary, public health, and environmental perspectives.
Conclusion
The emergence of HPAI H5N1 clade 2.3.4.4b in domestic dairy cattle and cats marks a significant development in the ongoing global influenza landscape. Understanding the virus's genomic characteristics, reassortment events, and mutation patterns is crucial for effective surveillance, outbreak management, and public health interventions to mitigate potential threats to human and animal populations. Continued research and collaboration across disciplines are essential in addressing the dynamic challenges posed by emerging infectious diseases like HPAI.
The study findings by researchers from Iowa State University-USA were published on a preprint server and are currently being peer reviewed.
Highly Pathogenic Avian Influenza A (H5N1) clade 2.3.4.4b Virus detected in dairy cattle
The global emergence of highly pathogenic avian influenza (HPAI) A (H5N1) clade 2.3.4.4b viruses poses a significant global public health threat. Until March 2024, no outbreaks of this virus clade had occurred in domestic cattle. We genetically characterize HPAI viruses from dairy cattle showing...
www.biorxiv.org
For the latest H5N1 News, keep on logging to Thailand Medical News.
Read Also:
https://www.thailandmedical.news/ne...ave-already-been-infected-with-the-new-strain
https://www.thailandmedical.news/ne...uza-do-not-work-against-the-new-h5n1-variants
Heliobas Disciple
TB Fanatic
(fair use applies)
Herbs And Phytochemicals: Study Shows That Zingerone And Shogoal From Ginger Can Inhibit SARS-CoV-2
Nikhil Prasad Fact checked by:Thailand Medical News Team
Apr 18, 2024
The global health landscape has witnessed a paradigm shift towards natural remedies in combating various diseases, particularly viral infections. The ongoing COVID-19 pandemic has highlighted the need for effective treatments derived from botanical sources. Among these natural compounds, ginger, with its rich array of bioactive components, has garnered significant attention for its potential in inhibiting SARS-CoV-2, the virus responsible for COVID-19.
Unlocking Ginger's Bioactive Arsenal
Ginger (Zingiber officinale), a well-known herbal plant, contains a myriad of bioactive compounds that contribute to its therapeutic properties. Notably, zingerone and shogaol, derived from gingerols, have emerged as key players in the realm of natural medicine. Zingerone, a major flavor component in ginger, exhibits antioxidant, anti-inflammatory, and immunomodulatory effects. Its multifaceted pharmacological profile positions it as a promising candidate for antiviral interventions. Shogaols, produced through the dehydration of gingerols, share similar properties and have been studied for their potential in combating various diseases, including cancer.
The Urgent Need for SARS-CoV-2 Inhibitors
The emergence of SARS-CoV-2 and the subsequent COVID-19 pandemic have underscored the critical need for effective antiviral agents. The spike (S) protein of SARS-CoV-2 plays a pivotal role in viral entry and replication, making it a prime target for therapeutic interventions. Inhibiting the interaction between the viral spike protein and host cell receptors is a key strategy in mitigating viral infectivity and spread.
Computational Insights: Evaluating Zingerone and Shogaol as Inhibitors
In a recent Herbs And Phytochemicals study conducted by researchers from Iraq, computational tools were employed to investigate the inhibitory potential of zingerone and shogaol against SARS-CoV-2. Utilizing advanced software such as Gaussian 09, UCA FUKUI, MGL implement, DSV, and LigPlus, the researchers delved into the molecular interactions between these natural compounds and the viral spike proteins.
Molecular Orbital Analysis: Unveiling Activity and Reactivity Patterns
The study employed molecular orbital analysis to elucidate the activity and reactivity patterns of zingerone and shogaol. Physical properties such as ELUMO (energy of the lowest unoccupied molecular orbital), energy gap, electronegativity, hardness, and dipole moment were assessed. These parameters provided insights into the compounds' electronic structure, which in turn influenced their potential as inhibitors.
The Fukui function and Dual Descriptor were instrumental in identifying the most active sites within zingerone and shogaol molecules. These computational techniques allowed researchers to predict the electron-accepting and electron-donating capabilities of specific atoms within the compounds, shedding light on their potential mechanis ms of action against SARS-CoV-2.
Interacting with Viral Spikes: Mechanistic Insights into Inhibition
Docking simulations were conducted to explore how zingerone and shogaol interacted with the viral spike proteins. The simulations revealed that zingerone and shogoal targeted key regions on the spike proteins, hindering the virus's ability to bind to host cell receptors. The binding energies calculated during the docking process provided quantitative measures of the strength of these interactions, with lower binding energies indicating stronger affinity and potential inhibitory activity.
Molecular Dynamics: Unraveling the Bonding Patterns
Detailed analyses of the interactions between zingerone, shogaol, and the amino acids comprising the viral spike proteins provided further insights into the mechanisms of inhibition. Hydrogen bonding, alkyl interactions, and pi-alkyl interactions were identified as critical mechanisms through which these compounds exerted their inhibitory effects. The specificity of these interactions highlighted the potential of zingerone and shogaol to disrupt essential viral processes and prevent viral entry and replication.
Conclusion: Harnessing Ginger's Power Against COVID-19
In conclusion, the study underscored the potential of zingerone and shogaol from ginger as inhibitors of SARS-CoV-2. Their ability to interact with viral spike proteins and disrupt key binding processes signifies a promising avenue for developing novel antiviral therapies. Further experimental validation, including in vitro and in vivo studies, is essential to leverage the full therapeutic potential of these natural compounds in combating COVID-19 and other viral infections. The findings of this study contribute to the growing body of evidence supporting the use of natural remedies in the fight against emerging viral threats.
The study findings were published in the peer reviewed journal: Parasite Epidemiology and Control (Elsevier).
[...]
Herbs And Phytochemicals: Study Shows That Zingerone And Shogoal From Ginger Can Inhibit SARS-CoV-2
Nikhil Prasad Fact checked by:Thailand Medical News Team
Apr 18, 2024
The global health landscape has witnessed a paradigm shift towards natural remedies in combating various diseases, particularly viral infections. The ongoing COVID-19 pandemic has highlighted the need for effective treatments derived from botanical sources. Among these natural compounds, ginger, with its rich array of bioactive components, has garnered significant attention for its potential in inhibiting SARS-CoV-2, the virus responsible for COVID-19.
Unlocking Ginger's Bioactive Arsenal
Ginger (Zingiber officinale), a well-known herbal plant, contains a myriad of bioactive compounds that contribute to its therapeutic properties. Notably, zingerone and shogaol, derived from gingerols, have emerged as key players in the realm of natural medicine. Zingerone, a major flavor component in ginger, exhibits antioxidant, anti-inflammatory, and immunomodulatory effects. Its multifaceted pharmacological profile positions it as a promising candidate for antiviral interventions. Shogaols, produced through the dehydration of gingerols, share similar properties and have been studied for their potential in combating various diseases, including cancer.
The Urgent Need for SARS-CoV-2 Inhibitors
The emergence of SARS-CoV-2 and the subsequent COVID-19 pandemic have underscored the critical need for effective antiviral agents. The spike (S) protein of SARS-CoV-2 plays a pivotal role in viral entry and replication, making it a prime target for therapeutic interventions. Inhibiting the interaction between the viral spike protein and host cell receptors is a key strategy in mitigating viral infectivity and spread.
Computational Insights: Evaluating Zingerone and Shogaol as Inhibitors
In a recent Herbs And Phytochemicals study conducted by researchers from Iraq, computational tools were employed to investigate the inhibitory potential of zingerone and shogaol against SARS-CoV-2. Utilizing advanced software such as Gaussian 09, UCA FUKUI, MGL implement, DSV, and LigPlus, the researchers delved into the molecular interactions between these natural compounds and the viral spike proteins.
Molecular Orbital Analysis: Unveiling Activity and Reactivity Patterns
The study employed molecular orbital analysis to elucidate the activity and reactivity patterns of zingerone and shogaol. Physical properties such as ELUMO (energy of the lowest unoccupied molecular orbital), energy gap, electronegativity, hardness, and dipole moment were assessed. These parameters provided insights into the compounds' electronic structure, which in turn influenced their potential as inhibitors.
The Fukui function and Dual Descriptor were instrumental in identifying the most active sites within zingerone and shogaol molecules. These computational techniques allowed researchers to predict the electron-accepting and electron-donating capabilities of specific atoms within the compounds, shedding light on their potential mechanis ms of action against SARS-CoV-2.
Interacting with Viral Spikes: Mechanistic Insights into Inhibition
Docking simulations were conducted to explore how zingerone and shogaol interacted with the viral spike proteins. The simulations revealed that zingerone and shogoal targeted key regions on the spike proteins, hindering the virus's ability to bind to host cell receptors. The binding energies calculated during the docking process provided quantitative measures of the strength of these interactions, with lower binding energies indicating stronger affinity and potential inhibitory activity.
Molecular Dynamics: Unraveling the Bonding Patterns
Detailed analyses of the interactions between zingerone, shogaol, and the amino acids comprising the viral spike proteins provided further insights into the mechanisms of inhibition. Hydrogen bonding, alkyl interactions, and pi-alkyl interactions were identified as critical mechanisms through which these compounds exerted their inhibitory effects. The specificity of these interactions highlighted the potential of zingerone and shogaol to disrupt essential viral processes and prevent viral entry and replication.
Conclusion: Harnessing Ginger's Power Against COVID-19
In conclusion, the study underscored the potential of zingerone and shogaol from ginger as inhibitors of SARS-CoV-2. Their ability to interact with viral spike proteins and disrupt key binding processes signifies a promising avenue for developing novel antiviral therapies. Further experimental validation, including in vitro and in vivo studies, is essential to leverage the full therapeutic potential of these natural compounds in combating COVID-19 and other viral infections. The findings of this study contribute to the growing body of evidence supporting the use of natural remedies in the fight against emerging viral threats.
The study findings were published in the peer reviewed journal: Parasite Epidemiology and Control (Elsevier).
[...]